


The role of long non-coding RNAs in breast cancer drug resistance: a comprehensive review
 0
0 Abstract
Breast cancer (BC) remains a major contributor to cancer-related morbidity and mortality among women globally. While systemic therapies have significantly advanced, the emergence of drug resistance continues to hinder durable clinical benefit, leading to treatment failure and disease relapse. Long non-coding RNAs (lncRNAs) have gained recognition as key regulators in BC biology and therapeutic response, with growing evidence implicating them in resistance to chemotherapy, endocrine therapy, targeted agents, and immunotherapy. This review provides a detailed overview of the molecular mechanisms through which specific lncRNAs promote resistance, emphasizing their roles in modulating processes such as programmed cell death, epithelial-mesenchymal transition, and remodeling of the tumor microenvironment. We further highlight common regulatory axes - such as competing endogenous RNA networks and the phosphatidylinositol 3-kinase/protein kinase B pathway - that serve as converging points across distinct resistance mechanisms. In addition, we discuss recent progress in harnessing lncRNAs as liquid biopsy biomarkers and examine the key challenges and opportunities in translating lncRNA-based targets into therapeutic strategies. Together, these insights offer a foundation for future research aimed at overcoming drug resistance in BC through novel lncRNA-centered interventions.
Keywords
INTRODUCTION
Breast cancer (BC) remains the most common malignancy and a leading cause of cancer-related mortality among women worldwide, posing a tremendous clinical and public health challenge[1,2]. While the advent of personalized medicine, guided by molecular subtyping (including hormone receptor status and human epidermal growth factor receptor 2 (HER2) expression), has significantly improved patient outcomes, the acquisition of drug resistance remains the primary obstacle to curative treatment across all major therapeutic strategies[3,4]. This resistance leads to disease progression, metastasis, and recurrence, ultimately limiting the efficacy of chemotherapy, endocrine therapy, targeted therapy, and the rapidly developing field of immunotherapy[5,6].
To overcome this pervasive clinical challenge, there is an urgent need to elucidate the intricate molecular mechanisms underpinning acquired drug resistance and identify novel therapeutic targets. Recent advances in genomics have revealed that long non-coding RNAs (lncRNAs), defined as RNA transcripts longer than 200 nucleotides lacking significant protein-coding potential, constitute a crucial layer of gene regulation[7-9]. Initially dismissed as transcriptional "noise", lncRNAs are now established as central molecular arbiters involved in almost every aspect of cancer biology, including initiation, progression, metastasis, and, critically, therapeutic response[10-12].
In BC, lncRNAs execute their regulatory functions through diverse mechanisms, such as acting as competing endogenous RNAs (ceRNAs), scaffolds for protein complexes, modulators of chromatin structure, and regulators of immune cell infiltration[13-15]. Accumulating evidence demonstrates that lncRNAs are master regulators of key cellular processes associated with drug resistance, including Epithelial-Mesenchymal Transition (EMT), modulation of the Tumor Microenvironment (TME), and various forms of Programmed Cell Death (PCD)[16-19]. Notably, recent high-impact studies have begun to uncover their critical roles in mediating resistance to novel therapeutic approaches, such as Ferroptosis[20,21] and Cuproptosis[22,23].
Given the complexity and clinical urgency of drug resistance, a comprehensive and up-to-date synthesis of lncRNA-mediated resistance is essential. This review systematically dissects the known functions and molecular mechanisms of lncRNAs across all four major therapeutic modalities: chemotherapy, endocrine therapy, targeted therapy, and immunotherapy. Crucially, we integrate these findings to highlight convergent regulatory pathways and shared molecular mechanisms that underpin lncRNA-mediated drug evasion, providing a unified view that transcends individual drug types.
ELUCIDATION OF MOLECULAR MECHANISMS: LNCRNAS AND THEIR REGULATION OF DRUG RESISTANCE IN BREAST CANCER
The challenge of drug resistance remains a formidable obstacle in BC therapy, manifesting across various domains, including resistance to chemotherapy, endocrine therapy, targeted therapy, and immunotherapy. The intricacies of drug resistance in BC involve a plethora of interconnected pathways: augmented DNA repair capacity, suppression of apoptotic gene expression, aberrant regulation of apoptotic proteins, cell cycle perturbations, alterations in drug target molecules, and heightened activity of drug efflux transporters[24]. Traditional research has largely centered on protein-coding genes implicated in drug resistance mechanisms, such as members of the adenosine triphosphate-binding cassette (ABC) transporter family, proteins related to EMT, and proteins involved in oxidative stress response[25-27]. However, these protein-coding genes represent a relatively modest portion of the human genome and exhibit limited tissue specificity, posing challenges for direct therapeutic targeting[28].
In contrast, lncRNAs, due to their ability to modulate downstream effectors and their heightened tissue and tumor specificity, have emerged as entities of considerable interest in oncological research. Numerous lncRNAs have been identified as key players in the orchestration of multidrug resistance in BC, particularly through the microRNA (miRNA)-ABC transporter axis. For example, lncRNA growth arrest-specific 5 (GAS5) has been observed to diminish the expression of ABC transport proteins, thereby potentiating the antitumor efficacy of doxorubicin in BC[29]. The lncRNA/EMT regulatory axis has been shown to influence tumor progression, with lncRNA Small Nucleolar RNA Host Gene 7 (SNHG7) driving EMT and consequent trastuzumab resistance by downregulating miRNA-186[30]. Additionally, lncRNAs have been implicated in the modulation of oxidative stress within BC cells, further delineating their multifaceted role in drug resistance[31,32].
Among the well-studied lncRNAs, the telomerase RNA component (TERC) plays a fundamental role in maintaining telomere length by acting as both scaffold and RNA template within the telomerase complex[33]. In conjunction with telomerase reverse transcriptase (TERT), TERC ensures chromosomal stability and sustained replicative capacity - features often exploited by cancer cells[34]. Dysregulation or mutation of TERC and TERT has been linked to BC development, and their aberrant activation may also reduce tumor sensitivity to DNA-damaging agents[35,36]. These findings highlight the TERC/TERT axis as a critical regulatory node in BC biology and a potential contributor to therapeutic resistance[37].
Recent insights reveal that certain lncRNAs can be conveyed to drug-sensitive cells via exosomes, effectuating the transmission of drug-resistant phenotypes. These lncRNAs govern the expression of gene sequences, modulate signaling cascades, influence cell cycle distribution, and impact drug-induced apoptosis[38-41]. Among these mechanisms, the most crucial involve lncRNAs functioning as ceRNAs or via exosomal pathways[42]. As BC evolves, certain lncRNAs, acting as ceRNAs, sequester and bind an increasing number of miRNAs. This process mitigates miRNA interactions with their target genes, resulting in augmented expression of these targets and subsequent alterations in key biological traits of BC, including invasiveness, metastatic potential, and drug resistance[24,43].
In summary, the mechanisms by which lncRNAs confer drug resistance in BC are profoundly diverse, ranging from intrinsic cellular changes to extrinsic microenvironmental modulation. At the cellular level, lncRNAs are central to the maintenance of the stemness phenotype and the induction of EMT, which collectively empower cancer cells with enhanced survival and metastatic potential[44]. Mechanistically, lncRNAs often function as scaffolds or ceRNAs, thereby perturbing canonical signaling cascades. Crucially, research across various therapeutic resistance models has revealed convergent regulatory modules, with lncRNAs frequently targeting and activating core survival pathways, notably the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR)[45-49], Wingless/Integrated (WnT)/β-catenin[50-52], and nuclear factor κB (NF-κB) pathways[53-55]. Furthermore, their role extends beyond the cell, with lncRNA cargo within exosomes facilitating paracrine communication with the TME, enabling the transfer of resistance phenotypes to sensitive cells and polarizing immune cells towards a suppressive state[56-59]. This convergence on shared pathways underscores a fundamental principle: despite the varying primary targets of different therapies, the ultimate mechanisms of drug evasion orchestrated by lncRNAs often share common regulatory hubs. A unified understanding of these shared mechanisms is essential to inform the development of pan-resistance strategies that target lncRNA-mediated vulnerability.
THE INTERPLAY BETWEEN LNCRNAS AND CHEMOTHERAPEUTIC RESISTANCE IN BREAST CANCER
Chemotherapy is a cornerstone in the battle against BC, instrumental in curbing tumor proliferation and staving off recurrence[60]. For individuals diagnosed with early-stage BC, adjuvant chemotherapy post-surgery can significantly reduce the likelihood of disease relapse. In advanced stages, neoadjuvant chemotherapy can decrease tumor burden, transforming inoperable cases into surgical candidates. The advent of chemotherapy has markedly enhanced both the disease-free and overall survival rates for BC patients[61]. Therapeutic regimens often include agents such as anthracyclines, paclitaxel, and cisplatin. Despite these strides, chemoresistance poses a persistent barrier to the effective management of BC[5]. The advent of combination chemotherapy, an approach addressing resistance to single-agent therapies, has become a standard practice in BC treatment, gaining widespread adoption in clinical settings[62,63]. This strategy employs a multifaceted arsenal of chemotherapeutic drugs, each at varying dosages, to elevate the success rate of chemotherapy. However, simply increasing the dosage of chemotherapeutic agents does not guarantee the eradication of all cancer cells. The nonlinear dynamics of dose-response relationships contribute to the emergence of chemoresistance[64]. Understanding and countering this resistance is pivotal for optimizing therapeutic efficacy and improving patient outcomes.
Mechanisms underpinning paclitaxel resistance in breast cancer
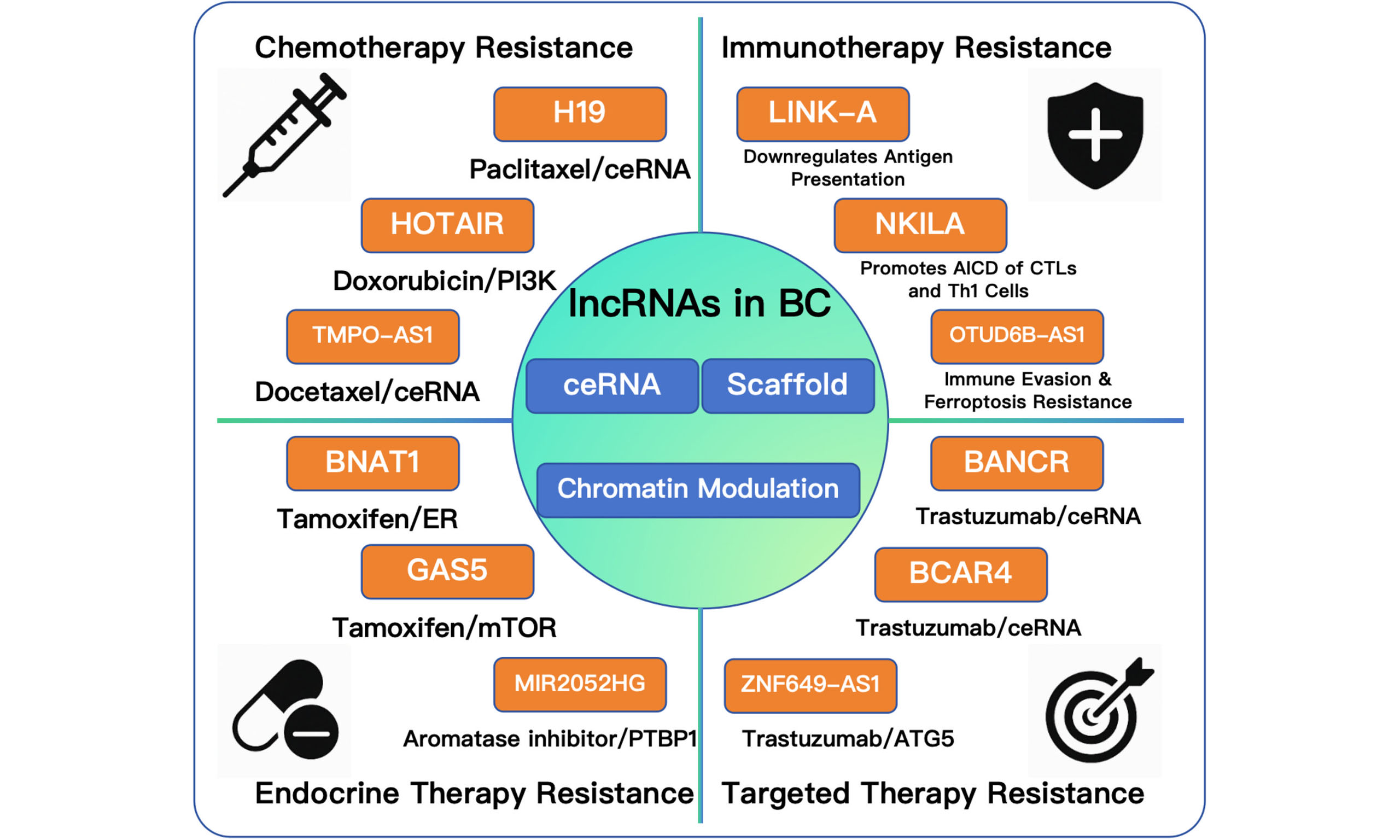
Paclitaxel remains a frontline chemotherapeutic agent in BC, primarily exerting its antitumor effect by inducing cell cycle arrest and promoting apoptosis. However, resistance to paclitaxel is common in clinical practice and significantly undermines treatment efficacy[65-67]. Recent studies have shed light on the role of lncRNAs in paclitaxel resistance within BC, many of which exhibit markedly increased expression levels. For instance, H19 Imprinted Maternally Expressed Transcript (H19), highly expressed in paclitaxel-resistant BC cell lines, functions as a ceRNA for miR-340-3p, consequently upregulating tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein zeta (YWHAZ) protein expression and fostering paclitaxel resistance[68]. Long intergenic non-coding RNA 00337 (LINC00337), with elevated presence in BC tissue, is implicated in enhancing malignancy and chemoresistance by stimulating alternatively activated macrophages (M2)-like macrophage-mediated tumor phenotypes, such as cell viability and migration, and EMT protein expression[69].
Moreover, research by Gu et al. pinpointed a long non-coding RNA entry from the NONCODE database (NONHSAT141924) as a significant associate of paclitaxel resistance in BC, with overexpression correlating with diminished survival rates in the Michigan Cancer Foundation-7 (MCF-7) BC cell line, potentially through modulation of the phosphorylated cAMP response element-binding protein (p-CREB)/B-cell lymphoma 2 (Bcl-2) apoptosis signaling pathway[70]. Contrarily, certain lncRNAs associated with paclitaxel resistance, including maternally expressed gene 3 (MEG3), have been found in reduced levels in BC tissues. The role of MEG3 may include the modulation of paclitaxel resistance and cancer cell malignancy via the miR-4513/probasin-like protein D (PBLD) axis[71]. Similarly, Zheng et al. observed a significant downregulation of GAS5 in paclitaxel-treated BC cells, with its overexpression bolstering cell sensitivity to paclitaxel and triggering apoptosis through the miR-378a-5p/suppressor of fused homolog (SUFU) signaling pathway[72].
These findings collectively underscore the complexity of lncRNA-mediated drug resistance mechanisms, presenting a nuanced landscape of gene expression and signaling pathways that orchestrate the cellular response to paclitaxel. This intricacy highlights the potential of lncRNAs as both biomarkers and therapeutic targets in overcoming chemoresistance and advancing BC treatment [Table 1].
LncRNAs and paclitaxel resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| Paclitaxel | AF178030.2 | Upregulation | TRPS1 | [73] |
| CASC2 | Upregulation | miR-18a-5p/CDK19 | [74] | |
| DDX11-AS1 | Upregulation | miRNA-497 | [75] | |
| FTH1P3 | Upregulation | miR-206/ABCB1 | [76] | |
| H19 | Upregulation | miR-340-3p/YWHAZ | [68] | |
| LINC00115 | Upregulation | SETDB1/PLK3/HIF1α | [77] | |
| LINC00160 | Upregulation | C/EBPβ/TFF3 | [78] | |
| LINC00337 | Upregulation | M2-like macrophages | [69] | |
| MALAT1 | Upregulation | miR-497-5p/SHOC2 | [79] | |
| NEAT1 | Upregulation | miR-133b | [80] | |
| NEAT1 | Upregulation | miR-23a-3p/FOXA1 | [81] | |
| NONHSAT141924 | Upregulation | p-CREB/Bcl-2 | [70] | |
| OTUD6B-AS1 | Upregulation | miR-26a-5p/MTDH | [82] | |
| PRKCQ-AS1 | Upregulation | miR-361-5p/PIK3C3 | [83] | |
| UCA1 | Upregulation | miR-613/CDK12 | [84] | |
| XIST | Upregulation | miR-let-7d-5p/ATG16L1 | [85] | |
| AC073284.4 | Downregulation | miR-18b-5p/DOCK4 | [86] | |
| GAS5 | Downregulation | miR-378a-5p/SUFU | [72] | |
| MEG3 | Downregulation | miR-4513/PBLD | [71] | |
| TPT1-AS1 | Downregulation | miR-3156-5p/caspase 2 | [87] |
Doxorubicin resistance mechanisms in breast cancer
Doxorubicin, an anthracycline antibiotic isolated from Streptomyces peucetius, is a principal chemotherapeutic agent employed against a spectrum of malignancies[88]. Its potent anticancer activity, characterized by the distinctive red chromophore, led to Food and Drug Administration (FDA) approval in 1974 for the treatment of malignancies such as lymphomas and leukemias and later expanded to include breast, ovarian, lung, and gastric cancers[89]. Doxorubicin acts primarily by intercalating DNA strands, impeding topoisomerase II activity, and consequently interrupting cellular DNA replication, leading to apoptosis in neoplastic cells[90]. Today, doxorubicin stands as a frontline agent in the adjuvant treatment of BC. Despite its efficacy, doxorubicin's clinical use is limited by significant cardiotoxic and neurotoxic side effects, although careful dose management may mitigate these risks[91].
In their research, Zhang et al. noted a pronounced upregulation of carbonyl reductase 3-antisense RNA 1 (CBR3-AS1) in BC tissues, which was strongly associated with poor patient prognosis. They proposed that CBR3-AS1 influences doxorubicin resistance in BC by modulating the miR-25-3p/mitogen-activated protein kinase kinase 4 (MEK4)/c-Jun N-terminal kinase 1 (JNK1) axis, with MEK4/JNK1 being the downstream effector of this axis. JNK1 silencing, in the context of CBR3-AS1 overexpression, appeared to enhance tissue sensitivity to doxorubicin[92]. Tang et al. identified that Dendritic Cell-Specific Transmembrane Protein 1 Antisense RNA 1 (DCST1-AS1) contributes to EMT through the Transforming Growth Factor β (TGF-β)/Smad signaling pathway by targeting annexin A1 (ANXA1), which may exacerbate doxorubicin resistance in triple-negative BC (TNBC) cells[93]. Luo et al. elucidated how plasmacytoma variant translocation 1 (PVT1) augments doxorubicin resistance by stabilizing nuclear factor erythroid 2-related factor 2 (Nrf2), disrupting the Kelch-like ECH-associated protein 1 (Keap1)-Nrf2 interaction[94]. Chen et al. reported that GAS5, underexpressed in doxorubicin-resistant BC cells, could, when overexpressed, impede the Wnt/β-catenin signaling pathway by interacting with miR-221-3p, reducing adenosine triphosphate-binding cassette sub-family B member 1 (ABCB1) expression and enhancing doxorubicin sensitivity[29]. Aini et al. integrated data from The Cancer Genome Atlas (TCGA) database and clinical samples to discover that small nucleolar RNA host gene 10 (SNHG10) was notably downregulated in TNBC relative to normal breast tissue. Their experimental findings indicated that SNHG10 negatively regulates miR-302b methylation, thereby impeding the development of doxorubicin resistance[95].
The comprehensive array of lncRNAs implicated in doxorubicin resistance in BC, detailed in Table 2, reflects the complexity of resistance mechanisms and underscores the potential of lncRNAs as therapeutic targets to circumvent chemoresistance and improve treatment efficacy.
LncRNAs and doxorubicin resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| Doxorubicin | BORG | Upregulation | NF-κB | [96] |
| CBR3-AS1 | Upregulation | miR-25-3p/MEK4/JNK1 | [92] | |
| DCST1-AS1 | Upregulation | ANXA1 | [93] | |
| DNAJC3-AS1 | Upregulation | miR-144 | [97] | |
| H19 | Upregulation | CUL4A/MDR1 | [98] | |
| HOTAIR | Upregulation | PI3K/AKT/mTOR | [45] | |
| LINC00518 | Upregulation | miR-199a/MRP1 | [99] | |
| LINC01977 | Upregulation | miR-212-3p/GOLM1 | [100] | |
| LOC645166 | Upregulation | NF-κB/GATA3 | [101] | |
| MALAT1 | Upregulation | miR-570-3p | [102] | |
| PVT1 | Upregulation | Nrf2 | [94] | |
| STMN1P2 | Upregulation | hnRNPU/EZH2/TRAF6 | [103] | |
| TUG1 | Upregulation | miR-9-5p/EIF5A2 | [104] | |
| XIST | Upregulation | miR-200c-3p/ANLN | [105] | |
| ZEB1-AS1 | Upregulation | miR-186-5p/ABCC1 | [106] | |
| GAS5 | Downregulation | miR-221-3p/DKK2 | [29] | |
| SNHG10 | Downregulation | miR-302b | [95] |
Docetaxel resistance mechanisms in breast cancer
Docetaxel, a taxane chemotherapy medication isolated from the needles of the European yew tree (Taxus baccata), has broad-spectrum efficacy against various malignancies, including breast, lung, prostate, and stomach cancers. Discovered by French scientists in the 1980s, docetaxel was found to be more potent than its taxane predecessor, paclitaxel. The FDA approved it in 1996 for advanced or metastatic BC treatment, and subsequently for additional cancer types[107]. Despite adverse effects such as alopecia, fatigue, and neutropenia, docetaxel has been integral to numerous chemotherapy protocols, significantly enhancing cancer patients' prognoses and quality of life[108].
While the pool of studies exploring lncRNAs associated with docetaxel resistance in BC remains relatively small, emerging research begins to illuminate this area (as shown in Table 3). For instance, Zhang et al. identified long intergenic non-protein coding RNA 461 (LINC00461) as overexpressed in BC tissues, correlating with unfavorable patient outcomes. Significantly, LINC00461 expression levels were elevated in docetaxel-resistant BC cell lines, where it was shown to foster cell proliferation, migration, and drug resistance by modulating miR-411-5p[109]. Moreover, Ning et al. found that thymopoietin antisense RNA 1 (TMPO-AS1) was markedly upregulated in docetaxel-resistant BC cells. Their study indicated that TMPO-AS1 downregulation enhanced the sensitivity of BC cells to docetaxel by interacting with miR-1179 and reducing tripartite motif-containing 37 (TRIM37) levels, thereby implicating the TMPO-AS1/miR-1179/TRIM37 axis as a key contributor to docetaxel resistance[110]. In addition, research by Li et al. pinpointed long intergenic non-protein coding RNA 667 (LINC00667)'s significant upregulation in exosomes from BC tissues and docetaxel-resistant cells, which contributed to decreased chemosensitivity by influencing the miR-200b-3p/Bcl-2 pathway[111].
LncRNAs and docetaxel resistance in breast cancer
These studies offer critical insights into docetaxel resistance mechanisms in BC, highlighting the importance of lncRNAs as mediators and potential therapeutic targets. This knowledge provides a foundation for devising new therapeutic strategies aimed at mitigating drug resistance and improving treatment outcomes in this challenging clinical scenario.
Epirubicin resistance mechanisms in breast cancer
Epirubicin, an anthracycline chemotherapeutic agent, is deployed against an array of cancers, including breast, ovarian, and gastrointestinal cancers. Pioneered by the Italian pharmaceutical entity Farmitalia Carlo Erba, this drug has secured regulatory approval across various nations, such as the United States, Canada, and Japan. Despite its clinical efficacy, epirubicin carries the burden of adverse effects, including myelosuppression, fatigue, alopecia, and nausea[112,113]. Efforts to mitigate these detrimental effects are underway, including its synergistic use with other chemotherapeutic compounds.
In a pivotal study by Yao et al., the correlation between epirubicin resistance and lncRNA expression in BC was thoroughly examined. The researchers established epirubicin-resistant BC cell sublines and applied high-throughput sequencing to pinpoint differentially expressed lncRNAs. Among them, a long non-coding RNA entry from the NONCODE database (NONHSAT101069) was markedly upregulated in clinical BC samples, BC cell lines, and epirubicin-resistant sublines. Subsequent analysis suggested that it serves as a ceRNA, sequestering miR-129-5p and consequently modulating the expression of twist-related protein 1 (Twist1), a key player in the pathogenesis of epirubicin resistance[114]. Additionally, Zheng et al. delved into the role of Sprouty receptor tyrosine kinase signaling antagonist 4-intronic transcript 1 (SPRY4-IT1) in BC, uncovering its significant overexpression and associations with increased lymph node metastasis and recurrence rates. Elevated levels of SPRY4-IT1 were found to confer chemoresistance to epirubicin in BC cell lines MCF-7 and MDA-MB-231[115]. Wang et al. reported that long intergenic non-protein coding RNA 5620 (LINC005620) was highly expressed in epirubicin-resistant TNBC cell lines and implicated in the facilitation of malignant TNBC progression and resistance to epirubicin by modulating integrin β 1 (ITGB1) expression[116].
Collectively, these investigations into lncRNAs present a comprehensive view of the molecular intricacies underpinning epirubicin resistance in BC, as illustrated in Table 4. The elucidation of these mechanisms holds the promise of steering the development of novel therapeutic strategies, targeting lncRNAs to surmount chemoresistance, and enhancing the treatment of this formidable disease.
5-fluorouracil resistance mechanisms in breast cancer
The 5-fluorouracil (5-FU), a pyrimidine analog used in chemotherapy, targets a variety of cancers, including those affecting the breast, colon, and stomach. Synthesized in the 1950s by Charles
Investigations into lncRNA contributions to 5-FU resistance in BC have revealed substantial insights.
These studies underscore the complexity of 5-FU chemoresistance mechanisms and the emergent role of lncRNAs as modulators of this process. They collectively present a nuanced understanding of BC pathology and treatment resistance, offering promising avenues for developing new therapeutic strategies to effectively counteract 5-FU resistance in BC, as detailed in Table 5.
LncRNAs and 5-fluorouracil resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| 5-Fluorouracil | CCAT2 | Upregulation | microRNA-145 | [122] |
| CDR1-AS | Upregulation | miR-7/CCNE1 | [124] | |
| MIAT | Upregulation | GRP78/OCT4/MIAT/AKT | [125] | |
| NEAT1 | Upregulation | miR-211/HMGA2 | [120] | |
| PRLB | Upregulation | miR-4766-5p/SIRT1 | [121] | |
| SNORD3A | Downregulation | miR-185-5p/UMPS | [123] |
Platinum resistance mechanisms in breast cancer
Platinum-based chemotherapeutics, named for their key component, platinum, are pivotal in treating various cancers, including ovarian, testicular, and lung cancers. Cisplatin, discovered in 1965 at Michigan State University, marked the advent of platinum drugs in cancer therapy. It was introduced in clinical practice in the late 1970s, establishing itself as a foundational chemotherapy agent for numerous malignancies. Successors such as carboplatin and oxaliplatin were developed during the 1980s, offering enhanced efficacy and a reduced side effect profile. Among these, cisplatin-based treatments are vital in managing BC[126], primarily through inducing DNA damage and mitochondrial apoptosis[127]. Nonetheless, the adverse effects, particularly nephrotoxicity and ototoxicity, persist as significant challenges[128-131]. Current research is aimed at augmenting their therapeutic index through drug combinations, novel compounds, and precision delivery techniques.
In the realm of lncRNA research, Zhou et al. pinpointed highly upregulated in liver cancer (HULC) as a pivotal regulator of insulin-like growth factor 1 receptor (IGF1R) in BC, with its upregulation promoting IGF1R expression and consequent activation of the PI3K/AKT pathway, enhancing cancer stemness and cisplatin resistance[132]. Su et al. delineated how differentiation antagonizing non-protein coding RNA (DANCR) interacts with Krüppel-like factor 5 (KLF5) in TNBC cells, modulating cyclin-dependent kinase inhibitor 1B (p27) transcription and influencing cisplatin chemoresistance[133]. The exploration of García-Venzor et al. into the role of lncRNA methionine adenosyl transferase 2 non-catalytic β subunit (lncMat2B) in BC revealed its association with increased survival under cisplatin treatment in MCF-7 cells, attributed to diminished DNA damage and reactive oxygen species production[134]. Tang et al. observed a marked reduction of taurine-upregulated gene 1 (TUG1) in TNBC tissues, unveiling its potential to modulate the Wnt signaling pathway through the miR-197/Nemo-like kinase (NLK) axis, thereby augmenting cisplatin sensitivity in TNBC cells[135]. Furthermore, Sheng et al. reported the downregulation of cardiac mesoderm-specific nuclear protein (CARMN) across different stages of breast malignancy, with its involvement in inhibiting DNA replication and amplifying cisplatin sensitivity via the miR143-3p/Minichromosome maintenance complex component 5 (MCM5) axis in TNBC[136].
These investigations elucidate the intricate role of lncRNAs in modulating platinum drug resistance in BC, providing a promising avenue for novel therapeutic strategies aimed at circumventing chemoresistance and enhancing treatment outcomes, as summarized in Table 6.
LncRNAs and cisplatin resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| Cisplatin | AFAP1-AS1 | Upregulation | miR-195/miR-545 | [137] |
| DANCR | Upregulation | KLF5/p27 | [133] | |
| HCP5 | Upregulation | PTEN | [138] | |
| HULC | Upregulation | IGF1R-PI3K-AKT | [132] | |
| LINC00511 | Upregulation | / | [139] | |
| LncMAT2B | Upregulation | DNA damage repair | [134] | |
| NEAT1 | Upregulation | microRNA-141-3p/KLF12 | [140] | |
| PART1 | Upregulation | / | [141] | |
| SNHG1 | Upregulation | miR-381 | [142] | |
| SNHG15 | Upregulation | miR-381 | [143] | |
| SNHG15 | Upregulation | miR-345-5p | [144] | |
| CARMN | Downregulation | miR143-3p | [136] | |
| MT1JP | Downregulation | miR-24-3p | [145] | |
| TUG1 | Downregulation | miR-197 | [135] |
THE ROLE OF LNCRNAS IN MODULATING ENDOCRINE THERAPY RESISTANCE IN BREAST CANCER
Endocrine therapy represents a cornerstone treatment for hormone-sensitive BC, aiming to curtail cancer cell proliferation by inhibiting hormone actions, notably estrogen, that promote tumor growth. This therapeutic approach spans the continuum of BC management, from neoadjuvant (pre-surgical) to adjuvant (post-surgical) therapy and is also crucial during salvage treatment for recurrent disease[146,147]. The pharmacological arsenal of endocrine therapy primarily comprises estrogen receptor antagonists, including tamoxifen and toremifene, which obstruct the oncogenic effects of estrogen on BC cells, and aromatase inhibitors, which diminish estrogen synthesis within the body[148]. The mechanisms underlying resistance to endocrine therapy are multifaceted and remain incompletely elucidated. However, emerging evidence highlights the significance of epigenetic modifications influencing estrogen levels, disruptions in estrogen signaling pathways, and the interplay between these pathways as pivotal factors contributing to endocrine therapy resistance in BC. These insights underscore the complexity of hormonal regulation and its impact on treatment efficacy[149,150]. Endocrine therapy is indispensable for managing hormone-sensitive BC, markedly lowering the risk of cancer recurrence and enhancing survival prospects. Nevertheless, the challenge of therapy resistance necessitates further investigation to decode the underlying mechanisms, with a particular focus on the contribution of lncRNAs. Understanding these aspects is crucial for developing innovative strategies to surmount resistance, thereby optimizing treatment outcomes for BC patients. The elucidation of lncRNA-mediated mechanisms offers promising avenues for future research aimed at enhancing the therapeutic arsenal against hormone-sensitive BC.
Tamoxifen resistance mechanisms in breast cancer
Tamoxifen (TAM), a pioneering selective estrogen receptor modulator (SERM), has been instrumental in both the treatment and prevention of BC by antagonizing estrogen's influence on breast tissues. Initially developed as a contraceptive in the 1960s, subsequent research unveiled its efficacy against BC, leading to FDA approval in 1973 for advanced cases and in 1998 for prophylaxis in high-risk individuals. As a cornerstone of endocrine therapy, especially for premenopausal women, tamoxifen significantly lowers BC risk by up to 50%, decreases mortality by about 30%, and reduces recurrence rates among survivors[147,151-153]. Enhancing the effectiveness of tamoxifen, including its use in combination therapies and the development of novel SERMs with minimized adverse effects, remains a focal point of ongoing research.
Recent studies have elucidated the role of lncRNAs in mediating tamoxifen resistance. Shi et al. discovered down-regulated in liver cancer 1 (DILA1), a lncRNA that aberrantly interacts with Cyclin D1 in tamoxifen-resistant cells. DILA1 suppression was observed to diminish Cyclin D1 levels, curb cancer cell proliferation, and re-sensitize cells to tamoxifen[154]. Wang et al. found a marked increase in H19 expression in tamoxifen-resistant cells and tissues, with its knockdown enhancing tamoxifen sensitivity by modulating autophagy via the S-adenosylhomocysteine hydrolase (SAHH)/DNA methyltransferase 3 beta (DNMT3B) axis[155]. Lin et al. pinpointed brain-derived neurotrophic factor antisense RNA (BDNF-AS), overexpressed due to Myocyte enhancer factor 2A (MEF2A)-regulated enhancers in endocrine-resistant BC and TNBC, which could drive resistance and tumor progression through the Ribonuclease Inhibitor 1 (RNH1)/Tripartite Motif-Containing 2 (TRIM21)/mTOR pathway[156]. Xue et al. reported the upregulation of HOX transcript antisense RNA (HOTAIR) in tamoxifen-resistant tissues, implicating its role in fostering ligand-independent estrogen receptor activity and consequent resistance[157]. Additionally, Li et al. identified significant upregulation of urothelial carcinoma-associated 1 (UCA1) in BC tissues, influencing tamoxifen resistance via the Enhancer of Zeste 2 Polycomb Repressive Complex 2 Subunit (EZH2)/Cyclin-Dependent Kinase Inhibitor 1 (p21) and PI3K/AKT pathways[158]. Furthermore, Shi et al. highlighted the underexpression of A disintegrin and metalloproteinase metallopeptidase with thrombospondin type 1 motif, 9-Antisense RNA 2 (ADAMTS9-AS2) in BC and tamoxifen-resistant cells, noting that reduced ADAMTS9-AS2 levels could attenuate phosphatase and tensin homolog (PTEN) expression and augment tamoxifen resistance by targeting miRNA-130a-5p[159].
These insights into lncRNA-mediated tamoxifen resistance underscore the complexity of hormonal therapy evasion in BC. While existing research has begun to unravel the intricacies of this resistance mechanism and the pivotal role of lncRNAs, further exploration is crucial for developing more effective therapeutic strategies to overcome tamoxifen resistance[160,161] (as shown in Table 7).
LncRNAs and tamoxifen resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| Tamoxifen | AGPG | Upregulation | PURα/E2F1 | [162] |
| ATXN8OS | Upregulation | miR-16-5p/VASP | [163] | |
| BDNF-AS | Upregulation | RNH1/TRIM21/mTOR | [156] | |
| BNAT1 | Upregulation | ER Signaling | [164] | |
| CYTOR | Upregulation | miR-125a-5p | [165] | |
| DILA1 | Upregulation | Cyclin D1 | [154] | |
| DLGAP1-AS2 | Upregulation | ER signaling | [166] | |
| DSCAM-AS1 | Upregulation | miR-137/EPS8 | [167] | |
| FOXD3-AS1 | Upregulation | miR-363 | [168] | |
| H19 | Upregulation | autophagy | [155] | |
| HNF1A-AS1 | Upregulation | miR-363/SERTAD3 | [169] | |
| HOTAIR | Upregulation | ER signaling | [157] | |
| HOTAIRM1 | Upregulation | HOXA1 | [170] | |
| IGF2-AS | Upregulation | / | [171] | |
| Lnc-DC | Upregulation | STAT3/Bcl-2 | [172] | |
| LINC00152 | Upregulation | TRPC1/ROS/ferroptosis | [173] | |
| LINC02568 | Upregulation | miR-1233-5p/ESR1 | [174] | |
| LINP1 | Upregulation | ER signaling | [175] | |
| LOL | Upregulation | let-7 | [176] | |
| MAFG-AS1 | Upregulation | miR-339-5p/CDK2 | [177] | |
| PRKCQ-AS1 | Upregulation | miR-200a-3p/MKP1 | [178] | |
| ROR | Upregulation | MAPK/ERK | [179] | |
| SNHG6 | Upregulation | miR-101 | [180] | |
| TTN-AS1 | Upregulation | miR-107/PI3K/AKT | [181] | |
| UCA1 | Upregulation | EZH2/p21 | [158] | |
| ADAMTS9-AS2 | Downregulation | miR-130a-5p/PTEN | [159] | |
| GAS5 | Downregulation | mTOR pathway | [49] |
Aromatase resistance mechanisms in breast cancer
Aromatase inhibitors constitute a cornerstone therapy for postmenopausal women with BC, targeting the enzyme aromatase to impede the conversion of androgens into estrogens, thereby diminishing estrogen levels. The genesis of aromatase inhibitors traces back to the 1960s with the development of aminoglutethimide, initially intended for Cushing's syndrome but later recognized for its capacity to lower estrogen levels in BC patients. Subsequent generations, including anastrozole and letrozole, emerged in the 1980s, showcasing superior efficacy over preceding treatments in minimizing BC recurrence risk and enhancing survival rates. Presently, these inhibitors are embraced as primary therapy for hormone receptor-positive BC in postmenopausal women[182-184]. Research endeavors continue to refine their clinical application and delineate predictive biomarkers to identify the subset of patients most likely to derive benefit.
Chen et al. unveiled that Deiodinase, Iodothyronine, Type III, Antisense RNA (DIO3OS) levels were heightened in tissues from patients exhibiting resistance to aromatase inhibitors. DIO3OS was found to foster metabolic adaptability in BC cells by activating the Polypyrimidine Tract Binding Protein 1 (PTBP1)/lactate dehydrogenase A (LDHA) glycolytic pathway, thereby ensuring cell survival under the estrogen-deprived conditions induced by aromatase inhibition[185]. Additionally, the retrospective analysis of 290 BC cases by Huang et al. underscored high Long Intergenic Non-Protein Coding RNA 309 (LINC00309) expression as a harbinger of adverse disease-free survival, establishing its association with reduced survival durations. High LINC00309 expression also correlated significantly with resistance to hormone therapies, including SERMs and aromatase inhibitors[186]. Moreover, the work by Ingle et al. highlighted the pivotal function of MIR2052 host gene (MIR2052HG) in sustaining estrogen receptor alpha (ERα) levels through the modulation of AKT/Forkhead Box O3 (FOXO3)-mediated estrogen receptor 1 (ESR1) transcription and curtailing ERα degradation via ubiquitination. The expression of MIR2052HG also correlated with recurrence risk in patients receiving aromatase inhibitors[187].
These investigations illuminate the critical involvement of lncRNAs in the resistance mechanisms to aromatase inhibitors in BC, underscoring the necessity for further study to unravel the intricate roles and mechanisms of these lncRNAs in the onset and progression of the disease, as delineated in Table 8.
LONG NON-CODING RNAS AND THEIR ROLE IN TARGETED THERAPY RESISTANCE IN BREAST CANCER
Targeted therapy represents a transformative approach in BC treatment, capitalizing on the selective targeting of molecules crucial for cancer growth and progression[188]. This strategy contrasts with conventional chemotherapy by offering enhanced efficacy and a reduced side effect profile due to its specificity for genetic aberrations driving tumorigenesis. By inhibiting signal transduction pathways essential for tumor cell survival, targeted therapies curtail tumor growth while mitigating systemic adverse effects, such as chemotherapy-associated skin ulceration and alopecia[189,190]. Determining a patient's eligibility for targeted therapy typically necessitates genetic testing to identify actionable mutations[191]. Advancements in the molecular understanding of BC have unveiled several targets, including HER2, Insulin-like Growth Factor Receptor (IGFR), FGFR, mTOR, and Cyclin-dependent kinase 4/6 (CDK4/6)[192], propelling the development of therapies such as trastuzumab for HER2-positive tumors, which constitute 18%-20% of BC cases[193]. Despite the effectiveness of targeted therapies, the response rate to trastuzumab in HER2-positive BC patients remains below 35%[194,195].
Trastuzumab, a monoclonal antibody developed in the 1990s by Genentech, revolutionized HER2-positive BC treatment by inhibiting HER2 signaling. FDA-approved in 1998 for metastatic and in 2006 for early-stage HER2-positive BC, trastuzumab significantly enhances survival rates, establishing it as a cornerstone in the management of this BC subtype. Ongoing research seeks to optimize trastuzumab's efficacy through combinational strategies and broaden its application to other HER2-expressing malignancies[196,197].
The intersection of lncRNAs with trastuzumab resistance has been a focal point of recent studies[184,198]. Dong et al. identified Small Nucleolar RNA Host Gene 14 (SNHG14) as a pro-tumorigenic lncRNA that also mediates trastuzumab resistance, acting through the modulation of Poly(A) Binding Protein Cytoplasmic 1 (PABPC1) expression via Histone H3 Lysine 27 (H3K27) acetylation[199]. Moreover, Zhu et al. elucidated the involvement of UCA1 in trastuzumab sensitivity modulation via the miR-18a/Yes-associated protein 1 (YAP1) axis[200].
These insights underscore the pivotal role of lncRNAs in shaping the landscape of targeted therapy resistance in BC, highlighting their potential as biomarkers and therapeutic targets. However, further research is imperative to decode the comprehensive mechanisms by which lncRNAs contribute to resistance and to integrate these findings into clinical practice.
While most current studies focus on linear lncRNAs, emerging evidence suggests that circular RNAs (circRNAs) may also participate in therapeutic resistance[201-203]. Notably, a recent investigation identified a circRNA named BISC that restores sensitivity to Bromodomain and Extra-Terminal domain proteins (BET) inhibitors in TNBC by blocking Insulin-Like Growth Factor 2 mRNA-Binding Protein 2 (IGF2BP2)-mediated translation of cellular MYC proto-oncogene (c-MYC)[204]. These findings not only broaden the spectrum of non-coding RNA involvement in drug resistance but also point to the potential of circRNA-based interventions in overcoming resistance to targeted therapies, as delineated in Table 9.
LncRNAs and trastuzumab resistance in breast cancer
| Drugs | LncRNAs | Expression | Genes and pathways | Reference |
| Trastuzumab | ATB | Upregulation | miR200c/ZEB1/ZNF-217 | [205] |
| BANCR | Upregulation | miR-34a-5p/HMGA1 | [206] | |
| BCAR4 | Upregulation | miR-665/YAP1/TGF-β | [207] | |
| H19 | Upregulation | / | [208] | |
| HOTAIR | Upregulation | TGF-β | [209] | |
| LINC00589 | Downregulation | miR-100/DLG5 | [210] | |
| LINC00969 | Upregulation | HuR/HER2 | [211] | |
| OIP5-AS1 | Upregulation | miR-381-3p/HMGB3 | [212] | |
| SNHG7 | Upregulation | miR-186 | [30] | |
| SNHG14 | Upregulation | H3K27/PABPC1 | [199] | |
| UCA1 | Upregulation | miR-18a/Yes | [200] | |
| ZNF649-AS1 | Upregulation | ATG5 | [213] | |
| ZNF649-AS1 | Upregulation | PRPF8/EXOC7-S | [214] | |
| GAS5 | Downregulation | miR-21/PTEN | [215] |
THE INFLUENCE OF LONG NON-CODING RNAS ON IMMUNOTHERAPY RESISTANCE IN BREAST CANCER
BC immunotherapy, an evolving frontier in cancer treatment, employs biological agents to enhance the ability of the immune system of patients to combat cancer. This therapy might be categorized as active, aiming to directly stimulate the immune response, or passive, involving the administration of immune cells or proteins. Advances in immunology have positioned immunotherapy as a viable adjunct to conventional treatments such as surgery, chemotherapy, radiotherapy, endocrine therapy, and targeted therapy, offering a potentially more precise and less detrimental modality for BC management. Initial studies underscored the immunomodulatory potential of agents such as Bacillus Calmette-Guérin (BCG) and various cytokines in BC[216]. Contemporary breakthroughs have ushered in novel immunotherapeutic approaches, including immune checkpoint blockade and T-cell therapy[217,218]. Despite BC's classification as a "cold" tumor, characterized by a subdued immune response, recent clinical trials, notably the IMpassion130 study (NCT02425891), have illuminated the efficacy of immunotherapy in treating TNBC[219], particularly when combined with atezolizumab and nab-paclitaxel for programmed death-ligand 1 (PD-L1)-positive TNBC, marking a significant advance in treatment paradigms[220].
However, the path to widespread immunotherapeutic success in BC is hindered by immune evasion and resistance mechanisms. Emerging research implicates lncRNAs in modulating immune responses, fostering an immunosuppressive tumor milieu, and contributing to resistance against immunotherapy.
Immuno-lncRNA signatures have been extensively developed to predict prognosis, immune status, and the sensitivity of BC patients to immune checkpoint inhibitors[221,222]. These lncRNAs regulate various PCD pathways, thereby influencing immune resistance. For example, signatures associated with Necroptosis[223,224] can predict the tumor immune microenvironment, while Cuproptosis-related ceRNA networks may imply mechanisms of endocrine or immunotherapy resistance[225]. Notably, lncRNA ovarian tumor domain-containing 6B antisense RNA 1 (OTUD6B-AS1) has been shown to exert a dual role, mediating both immune evasion and Ferroptosis resistance[20], highlighting its potential as a prognostic and therapeutic biomarker. Tumor cell metabolic reprogramming is closely linked to immune escape and resistance. The oncogenic lncRNA LINK-A (or LINC01139) is demonstrated to promote tumor immune evasion by downregulating cancer cell antigen presentation and inhibiting intrinsic tumor suppression[226,227]. Furthermore, the expression of lncRNA long intergenic non-protein coding RNA 2544 (LINC02544) drives immunotherapy resistance in TNBC through the miR-497-5p/cell cycle associated protein 1 (CAPRIN1) axis[228]. The influence of lncRNAs on the TME is also critical. For instance, PVT1 expressed by tumor-associated nonmyelinating Schwann cells (TASc) promotes the Kynurenine pathway and leads to tumor immune exclusion in pancreatic cancer[229], suggesting that lncRNAs regulate TME cell populations. Moreover, exosomal non-coding RNAs, including lncRNAs, are crucial mediators of drug and immunotherapy resistance in BC[230,231].
Notably, the LINK-A lncRNA has been linked to antigen loss and diminished tumor immunogenicity in metastatic BC[227]. NF-κB interacting long non-coding RNA (NKILA) expression was found to increase susceptibility of tumor-specific cytotoxic T lymphocytes and T helper type 1 (TH1) cells to activation-induced cell death (AICD) in a BC mouse model[232]. Additionally, Pei et al. demonstrated that targeting small nucleolar RNA host gene 1 (SNHG1) in BC cell lines could attenuate regulatory T cell (Treg) differentiation by upregulating miR-448 and downregulating indoleamine 2,3-dioxygenase (IDO) expression, potentially obstructing BC's immune escape[233].
These findings spotlight lncRNAs as pivotal regulators of the immune landscape in BC, influencing the efficacy of immunotherapy. The delineation of lncRNA-mediated pathways presents a promising avenue for overcoming immunotherapy resistance, paving the way for enhanced therapeutic strategies in BC management.
SUMMARY OF SHARED REGULATORY PATHWAYS
Despite the heterogeneity of therapeutic strategies in BC, accumulating evidence suggests that the mechanisms by which lncRNAs confer drug resistance often converge upon a limited set of conserved signaling pathways[19,234]. Elucidating these shared molecular nodes is essential for the development of unified therapeutic approaches capable of overcoming multidrug resistance.
A prominent mechanism involves the ceRNA network, whereby lncRNAs function as molecular decoys for specific miRNAs, thereby relieving suppression on downstream mRNAs involved in drug efflux, apoptosis, and EMT[235,236]. This regulatory axis has been widely implicated across multiple therapeutic contexts. For instance, H19 contributes to paclitaxel resistance through the miR-340-3p/YWHAZ pathway[237], while UCA1 mediates tamoxifen resistance by modulating the miR-18a/YAP1 axis[19].
Another frequently dysregulated node is the PI3K/AKT/mTOR pathway, which governs cell survival, proliferation, and metabolic reprogramming[238,239]. Multiple lncRNAs, including HOTAIR and MIR2052HG, have been shown to activate this cascade in the setting of doxorubicin, trastuzumab, and aromatase inhibitor resistance[240,241]. These findings emphasize the centrality of PI3K/AKT signaling as a conduit for lncRNA-driven therapeutic evasion.
The Wnt/β-catenin pathway has also emerged as a common target of lncRNA regulation, particularly in chemotherapy and immunotherapy resistance[242]. LncRNAs such as GAS5 and TUG1 influence this pathway by modulating apoptosis, stemness, and the expression of ABC transporters[19,243].
Moreover, NF-κB signaling, which plays a critical role in inflammation-mediated survival and resistance[244], is activated by lncRNAs such as BORG (BMP/OP-responsive gene, an lncRNA reported to activate NF-κB signaling and contribute to chemotherapy resistance) and (a long non-coding RNA annotated as a predicted genomic locus (LOC identifier)) (LOC645166), particularly in response to anthracycline-based chemotherapies[96,101]. These interactions support the role of lncRNAs in maintaining an inflammation-associated, pro-survival tumor phenotype.
In addition to intracellular signaling, exosome-mediated intercellular communication represents an emerging mechanism by which lncRNAs influence the TME[245,246]. Several lncRNAs, including H19, AGAP2-AS1, and UCA1, have been identified in exosomes derived from resistant cells, where they can transmit resistance traits to adjacent drug-sensitive cells[231]. This paracrine mechanism highlights the non-cell-autonomous role of lncRNAs in shaping resistance phenotypes and remodeling the TME.
Taken together, these convergent regulatory pathways - particularly the ceRNA network, PI3K/AKT, Wnt/β-catenin, NF-κB signaling, and exosomal transfer - represent key molecular hubs that integrate lncRNA-mediated resistance across diverse therapeutic contexts. Targeting these shared axes may offer a rational and potentially more effective strategy to overcome multidrug resistance in BC.
CURRENT CHALLENGES AND FUTURE PERSPECTIVES
Despite notable advances in delineating the roles of lncRNAs in BC drug resistance, several key obstacles must be addressed before these findings can be effectively translated into clinical application.
First, the functional characterization and mechanistic understanding of lncRNAs remain limited. While numerous lncRNAs have been associated with resistance phenotypes, only a small proportion have been mechanistically validated. Many exhibit limited sequence conservation, highly tissue- or context-specific expression, and complex subcellular localization patterns, complicating reproducibility across models and hindering definitive functional annotation.
Second, there is a scarcity of in vivo evidence and clinical correlation. Most available studies are confined to in vitro models or small patient cohorts, limiting their translational relevance. The absence of standardized preclinical animal models and the lack of prospective clinical datasets integrating lncRNA expression profiles pose significant barriers to establishing causality or evaluating predictive value in treatment response.
Third, nomenclatural inconsistency and incomplete genomic annotation continue to impede progress. Redundant naming, overlapping annotations, and the absence of curated entries in major databases contribute to confusion across studies and limit the utility of large-scale bioinformatics or integrative meta-analyses. Harmonizing lncRNA annotation frameworks will be essential to improve data standardization and cross-study comparability.
Fourth, therapeutic targeting of lncRNAs remains technically challenging. Although approaches such as antisense oligonucleotides (ASOs), small interfering RNAs (siRNAs), and CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats, referring to the genome-editing system mentioned in this Section)-based strategies have demonstrated promise in preclinical settings, their clinical applicability is hampered by issues of molecular instability, immunogenicity, poor tissue penetration, and off-target effects. Advanced delivery platforms - such as lipid nanoparticles, aptamer-conjugated carriers, or engineered exosomes - will be pivotal in enabling safe and effective lncRNA-targeted interventions.
Finally, the clinical implementation of lncRNA-based biomarkers requires rigorous validation. Although circulating or exosomal lncRNAs have shown potential as non-invasive indicators of drug resistance, their integration into routine diagnostics is constrained by technical variability in RNA isolation, quantification, and normalization. Multicenter, prospective trials incorporating standardized assays and clinically meaningful endpoints are urgently needed to establish their diagnostic, prognostic, or predictive utility.
Looking forward, future research should emphasize the systematic identification of pan-resistance lncRNAs, the construction of integrative regulatory networks, and the rational combination of lncRNA-targeted therapies with existing treatment regimens. Cross-disciplinary efforts bridging RNA biology, systems oncology, and translational medicine will be essential to overcome current limitations. Ultimately, a deeper mechanistic understanding of lncRNA-driven resistance may pave the way for more precise, durable, and personalized therapeutic strategies in BC.
CONCLUSIONS
lncRNAs have emerged as pivotal regulators of drug resistance in BC, influencing diverse biological processes and factors, including apoptosis, EMT, and the TME. Despite the heterogeneity of treatment modalities, mounting evidence indicates that many lncRNAs converge on common signaling axes - such as the ceRNA network, PI3K/AKT, Wnt/β-catenin, and NF-κB pathways - highlighting their potential as unifying therapeutic targets. While the clinical translation of lncRNA-based diagnostics and therapeutics remains challenging, continued efforts in mechanistic elucidation, in vivo validation, and delivery optimization hold promise for overcoming resistance and advancing precision oncology in BC.
DECLARATIONS
Authors’ contributions
Conceptualization, writing - original draft: Zeng Q, Lv B
Supervision, writing - review: Xu Y, Ma J
Validation, writing - editing: Ding C, Liang W
All authors reviewed the manuscript.
Availability of data and materials
Not applicable.
Financial support and sponsorship
None
Conflicts of interest
Xu Y is a Junior Editorial Board Member of Journal of Translational Genetics and Genomics. Xu Y was not involved in any part of the editorial process of this manuscript, including reviewer selection, manuscript handling, or decision-making, while the other authors have declared that they have no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2025.
REFERENCES
1. Bray F, Laversanne M, Sung H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024;74:229-63.
2. Giaquinto AN, Sung H, Newman LA, et al. Breast cancer statistics 2024. CA Cancer J Clin. 2024;74:477-95.
3. Perou CM, Sørlie T, Eisen MB, et al. Molecular portraits of human breast tumours. Nature. 2000;406:747-52.
4. Baselga J, Cortés J, Kim SB, et al. Pertuzumab plus trastuzumab plus docetaxel for metastatic breast cancer. N Engl J Med. 2012;366:109-19.
6. Karami Fath M, Azargoonjahromi A, Kiani A, et al. The role of epigenetic modifications in drug resistance and treatment of breast cancer. Cell Mol Biol Lett. 2022;27:52.
7. Ponting CP, Haerty W. Genome-wide analysis of human long noncoding RNAs: a provocative review. Annu Rev Genomics Hum Genet. 2022;23:153-72.
8. Shi K, Liu T, Fu H, Li W, Zheng X. Genome-wide analysis of lncRNA stability in human. PLoS Comput Biol. 2021;17:e1008918.
9. Mattick JS, Amaral PP, Carninci P, et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol. 2023;24:430-47.
10. Chen B, Dragomir MP, Yang C, Li Q, Horst D, Calin GA. Targeting non-coding RNAs to overcome cancer therapy resistance. Signal Transduct Target Ther. 2022;7:121.
11. Zhou W, Xu X, Cen Y, Chen J. The role of lncRNAs in the tumor microenvironment and immunotherapy of melanoma. Front Immunol. 2022;13:1085766.
12. Peng WX, Koirala P, Mo YY. LncRNA-mediated regulation of cell signaling in cancer. Oncogene. 2017;36:5661-7.
14. Chen LL. The biogenesis and emerging roles of circular RNAs. Nat Rev Mol Cell Biol. 2016;17:205-11.
15. Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and cancer: a new paradigm. Cancer Res. 2017;77:3965-81.
16. Kichi ZA, Soltani M, Rezaei M, Shirvani-Farsani Z, Rojhannezhad M. The emerging role of EMT-related lncRNAs in therapy resistance and their applications as biomarkers. Curr Med Chem. 2022;29:4574-601.
17. Pan G, Liu Y, Shang L, Zhou F, Yang S. EMT-associated microRNAs and their roles in cancer stemness and drug resistance. Cancer Commun. 2021;41:199-217.
18. Jiang N, Zhang X, Gu X, Li X, Shang L. Progress in understanding the role of lncRNA in programmed cell death. Cell Death Discov. 2021;7:30.
19. Ye P, Feng L, Shi S, Dong C. The mechanisms of lncRNA-mediated multidrug resistance and the clinical application prospects of lncRNAs in breast cancer. Cancers. 2022;14:2101.
20. Zhang JN, Yi ZL, Zhou XR, Liu SS, Liu H. Dual role of lncRNA OTUD6B-AS1 in immune evasion and ferroptosis resistance: a prognostic and therapeutic biomarker in breast cancer. Noncoding RNA Res. 2025;14:156-65.
21. Tong X, Yu Z, Xing J, et al. LncRNA HCP5-encoded protein regulates ferroptosis to promote the progression of triple-negative breast cancer. Cancers. 2023;15:1880.
22. Li C, Zhang Y. Construction and validation of a cuproptosis-related five-lncRNA signature for predicting prognosis, immune response and drug sensitivity in breast cancer. BMC Med Genom. 2023;16:158.
23. Wu JH, Cheng TC, Zhu B, Gao HY, Zheng L, Chen WX. Identification of cuproptosis-related gene SLC31A1 and upstream LncRNA-miRNA regulatory axis in breast cancer. Sci Rep. 2023;13:18390.
24. Du T, Shi Y, Xu S, Wan X, Sun H, Liu B. Long non-coding RNAs in drug resistance of breast cancer. Onco Targets Ther. 2020;13:7075-87.
25. Wang Y, Wang Y, Qin Z, et al. The role of non-coding RNAs in ABC transporters regulation and their clinical implications of multidrug resistance in cancer. Expert Opin Drug Metab Toxicol. 2021;17:291-306.
26. Blackley EF, Loi S. Targeting immune pathways in breast cancer: review of the prognostic utility of TILs in early stage triple negative breast cancer (TNBC). Breast. 2019;48 Suppl 1:S44-8.
27. Huynh MM, Pambid MR, Jayanthan A, Dorr A, Los G, Dunn SE. The dawn of targeted therapies for triple negative breast cancer (TNBC): a snapshot of investigational drugs in phase I and II trials. Expert Opin Investig Drugs. 2020;29:1199-208.
28. Djebali S, Davis CA, Merkel A, et al. Landscape of transcription in human cells. Nature. 2012;489:101-8.
29. Chen Z, Pan T, Jiang D, et al. The lncRNA-GAS5/miR-221-3p/DKK2 Axis modulates ABCB1-mediated adriamycin resistance of breast cancer via the wnt/β-catenin signaling pathway. Mol Ther Nucleic Acids. 2020;19:1434-48.
30. Zhang H, Zhang XY, Kang XN, Jin LJ, Wang ZY. LncRNA-SNHG7 enhances chemotherapy resistance and cell viability of breast cancer cells by regulating miR-186. Cancer Manag Res. 2020;12:10163-72.
31. Liu PF, Farooqi AA, Peng SY, et al. Regulatory effects of noncoding RNAs on the interplay of oxidative stress and autophagy in cancer malignancy and therapy. Semin Cancer Biol. 2022;83:269-82.
32. Kacso TP, Zahu R, Tirpe A, Paslari EV, Nuțu A, Berindan-Neagoe I. Reactive oxygen species and long non-coding RNAs, an unexpected crossroad in cancer cells. Int J Mol Sci. 2022;23:10133.
33. Baylie T, Jemal M, Baye G, et al. The role of telomere and telomerase in cancer and novel therapeutic target: narrative review. Front Oncol. 2025;15:1542930.
34. Dratwa M, Wysoczańska B, Łacina P, Kubik T, Bogunia-Kubik K. TERT-regulation and roles in cancer formation. Front Immunol. 2020;11:589929.
35. Lu L, Zhang C, Zhu G, et al. Telomerase expression and telomere length in breast cancer and their associations with adjuvant treatment and disease outcome. Breast Cancer Res. 2011;13:R56.
36. Lipinska N, Romaniuk A, Paszel-Jaworska A, Toton E, Kopczynski P, Rubis B. Telomerase and drug resistance in cancer. Cell Mol Life Sci. 2017;74:4121-32.
37. Cayuela ML, Flores JM, Blasco MA. The telomerase RNA component Terc is required for the tumour-promoting effects of Tert overexpression. EMBO Rep. 2005;6:268-74.
38. Hu W, Tan C, He Y, Zhang G, Xu Y, Tang J. Functional miRNAs in breast cancer drug resistance. Onco Targets Ther. 2018;11:1529-41.
39. Wang X, Pei X, Guo G, et al. Exosome-mediated transfer of long noncoding RNA H19 induces doxorubicin resistance in breast cancer. J Cell Physiol. 2020;235:6896-904.
40. Zheng Z, Chen M, Xing P, Yan X, Xie B. Increased expression of exosomal AGAP2-AS1 (AGAP2 antisense RNA 1) in breast cancer cells inhibits trastuzumab-induced cell cytotoxicity. Med Sci Monit. 2019;25:2211-20.
41. Xu CG, Yang MF, Ren YQ, Wu CH, Wang LQ. Exosomes mediated transfer of lncRNA UCA1 results in increased tamoxifen resistance in breast cancer cells. Eur Rev Med Pharmacol Sci. 2016;20:4362-8.
42. Kansara S, Pandey V, Lobie PE, Sethi G, Garg M, Pandey AK. Mechanistic involvement of long non-coding RNAs in oncotherapeutics resistance in triple-negative breast cancer. Cells. 2020;9:1511.
43. Veneziano D, Marceca GP, Di Bella S, Nigita G, Distefano R, Croce CM. Investigating miRNA-lncRNA interactions: computational tools and resources. In: Laganà A, editor. MicroRNA target identification. New York: Springer; 2019. pp. 251-77.
44. Guz M, Jeleniewicz W, Cybulski M. Interactions between circRNAs and miR-141 in cancer: from pathogenesis to diagnosis and therapy. Int J Mol Sci. 2023;24:11861.
45. Li Z, Qian J, Li J, Zhu C. Knockdown of lncRNA-HOTAIR downregulates the drug-resistance of breast cancer cells to doxorubicin via the PI3K/AKT/mTOR signaling pathway. Exp Ther Med. 2019;18:435-42.
46. Suicmez M, Namalir G, Konus M, Ozdil H. Silencing of lncRNA UCA1 reverses doxorubicin resistance of breast cancer through inhibiting PI3K/AKT/mTOR signaling pathway. ChemistrySelect. 2024;9:e202400819.
47. Lei C, Li S, Fan Y, et al. LncRNA DUXAP8 induces breast cancer radioresistance by modulating the PI3K/AKT/mTOR pathway and the EZH2-E-cadherin/RHOB pathway. Cancer Biol Ther. 2022;23:1-13.
48. Sadeghalvad M, Mansouri K, Mohammadi-Motlagh HR, et al. Long non-coding RNA HOTAIR induces the PI3K/AKT/mTOR signaling pathway in breast cancer cells. Rev Assoc Med Bras. 2022;68:456-62.
49. Jiang Y, Qian T, Li S, Xie Y, Tao M. Metformin reverses tamoxifen resistance through the lncRNA GAS5-medicated mTOR pathway in breast cancer. Ann Transl Med. 2022;10:366.
50. Xiao C, Wu CH, Hu HZ. LncRNA UCA1 promotes epithelial-mesenchymal transition (EMT) of breast cancer cells via enhancing Wnt/β-catenin signaling pathway. Eur Rev Med Pharmacol Sci. 2016;20:2819-24.
51. Liu H, Wang G, Yang L, Qu J, Yang Z, Zhou X. Knockdown of long non-coding RNA UCA1 increases the tamoxifen sensitivity of breast cancer cells through inhibition of Wnt/β-catenin pathway. PLoS One. 2016;11:e0168406.
52. Huang H, Jin H, Lei R, et al. lncRNA-WAL promotes triple-negative breast cancer aggression by inducing β-catenin nuclear translocation. Mol Cancer Res. 2024;22:1036-50.
53. Dashti S, Ghafouri-Fard S, Esfandi F, Oskooei VK, Arsang-Jang S, Taheri M. Expression analysis of NF-κB interacting long noncoding RNAs in breast cancer. Exp Mol Pathol. 2020;112:104359.
54. Liu B, Sun L, Liu Q, et al. A cytoplasmic NF-κB interacting long noncoding RNA blocks IκB phosphorylation and suppresses breast cancer metastasis. Cancer Cell. 2015;27:370-81.
55. Xu Y, Ren W, Li Q, et al. LncRNA Uc003xsl.1-mediated activation of the NFκB/IL8 axis promotes progression of triple-negative breast cancer. Cancer Res. 2022;82:556-70.
56. Zhang D, Ding X, Peng M. LncRNA SNHG14 accelerates breast cancer progression through sponging miR-543 and regulating KLF7 expression. Arch Gynecol Obstet. 2022;305:1507-16.
57. Xie SD, Qin C, Jin LD, et al. Long noncoding RNA SNHG14 promotes breast cancer cell proliferation and invasion via sponging miR-193a-3p. Eur Rev Med Pharmacol Sci. 2019;23:2461-8.
58. Zhang Y, Li Z, Chen M, et al. lncRNA TCL6 correlates with immune cell infiltration and indicates worse survival in breast cancer. Breast Cancer. 2020;27:573-85.
59. Adewunmi O, Shen Y, Zhang XH, Rosen JM. Targeted inhibition of lncRNA malat1 alters the tumor immune microenvironment in preclinical syngeneic mouse models of triple-negative breast cancer. Cancer Immunol Res. 2023;11:1462-79.
60. Desai P, Thumma NJ, Wagh PR, et al. Cancer chemoprevention using nanotechnology-based approaches. Front Pharmacol. 2020;11:323.
61. Zhang X, Xie K, Zhou H, et al. Role of non-coding RNAs and RNA modifiers in cancer therapy resistance. Mol Cancer. 2020;19:47.
62. Trayes KP, Cokenakes SEH. Breast cancer treatment. Am Fam Phys. 2021;104:171-8. Available from: https://www.aafp.org/pubs/afp/issues/2021/0800/p171.html [Last accessed on 4 Dec 2025].
63. Loibl S, Poortmans P, Morrow M, Denkert C, Curigliano G. Breast cancer. Lancet. 2021;397:1750-69.
64. Breast Cancer Trialists' Collaborative Group (EBCTCG). Increasing the dose intensity of chemotherapy by more frequent administration or sequential scheduling: a patient-level meta-analysis of 37 298 women with early breast cancer in 26 randomised trials. Lancet. 2019;393:1440-52.
65. Ahmed Khalil A, Rauf A, Alhumaydhi FA, et al. Recent developments and anticancer therapeutics of paclitaxel: an update. Curr Pharm Des. 2022;28:3363-73.
66. Jordan MA, Wilson L. Microtubules as a target for anticancer drugs. Nat Rev Cancer. 2004;4:253-65.
67. Newman DJ, Cragg GM, Snader KM. Natural products as sources of new drugs over the period 1981-2002. J Nat Prod. 2003;66:1022-37.
68. Yan L, Yang S, Yue CX, et al. Long noncoding RNA H19 acts as a miR-340-3p sponge to promote epithelial-mesenchymal transition by regulating YWHAZ expression in paclitaxel-resistant breast cancer cells. Environ Toxicol. 2020;35:1015-28.
69. Xing Z, Zhang M, Liu J, Liu G, Feng K, Wang X. LINC00337 induces tumor development and chemoresistance to paclitaxel of breast cancer by recruiting M2 tumor-associated macrophages. Mol Immunol. 2021;138:1-9.
70. Gu M, Zheng W, Zhang M, et al. LncRNA NONHSAT141924 promotes paclitaxel chemotherapy resistance through p-CREB/Bcl-2 apoptosis signaling pathway in breast cancer. J Cancer. 2020;11:3645-54.
71. Zhu M, Wang F, Mi H, et al. Long noncoding RNA MEG3 suppresses cell proliferation, migration and invasion, induces apoptosis and paclitaxel-resistance via miR-4513/PBLD axis in breast cancer cells. Cell Cycle. 2020;19:3277-88.
72. Zheng S, Li M, Miao K, Xu H. lncRNA GAS5-promoted apoptosis in triple-negative breast cancer by targeting miR-378a-5p/SUFU signaling. J Cell Biochem. 2020;121:2225-35.
73. Zhao T, Zhang T, Zhang Y, Zhou B, Lu X. Paclitaxel resistance modulated by the interaction between TRPS1 and AF178030.2 in triple-negative breast cancer. Evid Based Complement Alternat Med. 2022;2022:6019975.
74. Zheng P, Dong L, Zhang B, et al. Long noncoding RNA CASC2 promotes paclitaxel resistance in breast cancer through regulation of miR-18a-5p/CDK19. Histochem Cell Biol. 2019;152:281-91.
75. Liang M, Zhu B, Wang M, Jin J. Knockdown of long non-coding RNA DDX11-AS1 inhibits the proliferation, migration and paclitaxel resistance of breast cancer cells by upregulating microRNA-497 expression. Mol Med Rep. 2022;25:123.
76. Wang R, Zhang T, Yang Z, Jiang C, Seng J. Long non-coding RNA FTH1P3 activates paclitaxel resistance in breast cancer through miR-206/ABCB1. J Cell Mol Med. 2018;22:4068-75.
77. Luo F, Zhang M, Sun B, et al. LINC00115 promotes chemoresistant breast cancer stem-like cell stemness and metastasis through SETDB1/PLK3/HIF1α signaling. Mol Cancer. 2024;23:60.
78. Wu H, Gu J, Zhou D, et al. LINC00160 mediated paclitaxel-and doxorubicin-resistance in breast cancer cells by regulating TFF3 via transcription factor C/EBPβ. J Cell Mol Med. 2020;24:8589-602.
79. Shi C, Ren S, Zhao X, Li Q. lncRNA MALAT1 regulates the resistance of breast cancer cells to paclitaxel via the miR-497-5p/SHOC2 axis. Pharmacogenomics. 2022;23:973-85.
80. Wei X, Tao S, Mao H, et al. Exosomal lncRNA NEAT1 induces paclitaxel resistance in breast cancer cells and promotes cell migration by targeting miR-133b. Gene. 2023;860:147230.
81. Zhu L, Wang F, Fan W, Jin Z, Teng C, Zhang J. lncRNA NEAT1 promotes the Taxol resistance of breast cancer via sponging the miR-23a-3p-FOXA1 axis. Acta Biochim Biophys Sin. 2021;53:1198-206.
82. Li PP, Li RG, Huang YQ, Lu JP, Zhang WJ, Wang ZY. LncRNA OTUD6B-AS1 promotes paclitaxel resistance in triple negative breast cancer by regulation of miR-26a-5p/MTDH pathway-mediated autophagy and genomic instability. Aging. 2021;13:24171-91.
83. Zheng S, Fu W, Huang Q, et al. LncRNA PRKCQ-AS1 regulates paclitaxel resistance in triple-negative breast cancer cells through miR-361-5p/PIK3C3 mediated autophagy. Clin Exp Pharmacol Physiol. 2023;50:431-42.
84. Liu C, Jiang F, Zhang X, Xu X. Long non-coding RNA UCA1 modulates paclitaxel resistance in breast cancer via miR-613/CDK12 axis. Cancer Manag Res. 2020;12:2777-88.
85. Wang Y, Pei W, Yang Y, et al. Inhibition of XIST restrains paclitaxel resistance in breast cancer cells by targeting hsa-let-7d-5p/ATG16L1 through regulation of autophagy. Cell Signal. 2025;127:111534.
86. Wang YY, Yan L, Yang S, et al. Long noncoding RNA AC073284.4 suppresses epithelial-mesenchymal transition by sponging miR-18b-5p in paclitaxel-resistant breast cancer cells. J Cell Physiol. 2019;234:23202-15.
87. Huang Y, Zheng Y, Shao X, Shi L, Li G, Huang P. Long non-coding RNA TPT1-AS1 sensitizes breast cancer cell to paclitaxel and inhibits cell proliferation by miR-3156-5p/caspase 2 axis. Hum Cell. 2021;34:1244-54.
88. Arcamone F, Cassinelli G, Fantini G, et al. Adriamycin, 14-Hydroxydaunomycin, a new antitumor antibiotic fromS. peucetius var.caesius. Biotechnol Bioeng. 2000;67:704-13.
89. Thorn CF, Oshiro C, Marsh S, et al. Doxorubicin pathways: pharmacodynamics and adverse effects. Pharmacogenet Genom. 2011;21:440-6.
90. Renu K, V G A, P B TP, Arunachalam S. Molecular mechanism of doxorubicin-induced cardiomyopathy - an update. Eur J Pharmacol. 2018;818:241-53.
91. Lefrak EA, Piťha J, Rosenheim S, Gottlieb JA. A clinicopathologic analysis of adriamycin cardiotoxicity. Cancer. 1973;32:302-14.
92. Zhang M, Wang Y, Jiang L, et al. LncRNA CBR3-AS1 regulates of breast cancer drug sensitivity as a competing endogenous RNA through the JNK1/MEK4-mediated MAPK signal pathway. J Exp Clin Cancer Res. 2021;40:41.
93. Tang L, Chen Y, Chen H, et al. DCST1-AS1 promotes TGF-β-induced epithelial-mesenchymal transition and enhances chemoresistance in triple-negative breast cancer cells via ANXA1. Front Oncol. 2020;10:280.
94. Luo Y, Zhang W, Xu L, Chen Y, Xu Y, Yuan L. Long non-coding RNA PVT1 regulates the resistance of the breast cancer cell line MDA-MB-231 to doxorubicin via Nrf2. Technol Cancer Res Treat. 2020;19:1533033820980763.
95. Aini S, Bolati S, Ding W, et al. LncRNA SNHG10 suppresses the development of doxorubicin resistance by downregulating miR-302b in triple-negative breast cancer. Bioengineered. 2022;13:11430-9.
96. Gooding AJ, Zhang B, Gunawardane L, Beard A, Valadkhan S, Schiemann WP. The lncRNA BORG facilitates the survival and chemoresistance of triple-negative breast cancers. Oncogene. 2019;38:2020-41.
97. Ren R, Yuan Z, Xu Z. miRNA-144 targeting DNAJC3-AS1 reverses the resistance of the breast cancer cell line Michigan Cancer Foundation-7 to doxorubicin. Bioengineered. 2021;12:9885-92.
98. Zhu QN, Wang G, Guo Y, et al. LncRNA H19 is a major mediator of doxorubicin chemoresistance in breast cancer cells through a cullin4A-MDR1 pathway. Oncotarget. 2017;8:91990-2003.
99. Chang L, Hu Z, Zhou Z, Zhang H. Linc00518 contributes to multidrug resistance through regulating the MiR-199a/MRP1 axis in breast cancer. Cell Physiol Biochem. 2018;48:16-28.
100. Li Z, Li Y, Wang X, et al. LINC01977 promotes breast cancer progression and chemoresistance to doxorubicin by targeting miR-212-3p/GOLM1 axis. Front Oncol. 2021;11:657094.
101. Zheng R, Jia J, Guan L, et al. Long noncoding RNA lnc-LOC645166 promotes adriamycin resistance via NF-κB/GATA3 axis in breast cancer. Aging. 2020;12:8893-912.
102. Yue X, Wu WY, Dong M, Guo M. LncRNA MALAT1 promotes breast cancer progression and doxorubicin resistance via regulating miR-570-3p. Biomed J. 2021;44:S296-304.
103. Jin YP, Xu BJ, Zhang XF, et al. Long non-coding RNA STMN1P2 promotes breast cancer doxorubicin resistance by inhibiting pyroptosis through the hnRNPU-EZH2-TARF6-MALT1-caspase-1 pathway. Acta Pharmacol Sin. 2025:1-15.
104. Wang S, Cheng M, Zheng X, et al. Interactions between lncRNA TUG1 and miR-9-5p modulate the resistance of breast cancer cells to doxorubicin by regulating eIF5A2. Onco Targets Ther. 2020;13:13159-70.
105. Zhang M, Wang F, Xiang Z, Huang T, Zhou WB. LncRNA XIST promotes chemoresistance of breast cancer cells to doxorubicin by sponging miR-200c-3p to upregulate ANLN. Clin Exp Pharmacol Physiol. 2020;47:1464-72.
106. Lu Q, Chen W, Ji Y, Liu Y, Xue X. Ursolic acid enhances cytotoxicity of doxorubicin-resistant triple-negative breast cancer cells via ZEB1-AS1/miR-186-5p/ABCC1 axis. Cancer Biother Radiopharm. 2022;37:673-83.
108. Minckwitz G. Docetaxel/anthracycline combinations for breast cancer treatment. Expert Opin Pharmacother. 2007;8:485-95.
109. Zhang C, Wang J, Zhang J, Qu H, Tang X. LINC00461 overexpression can induce docetaxel resistance in breast cancer by interacting with miR-411-5p. Onco Targets Ther. 2020;13:5551-62.
110. Ning X, Zhao J, He F, Yuan Y, Li B, Ruan J. Long non-coding RNA TMPO-AS1 facilitates chemoresistance and invasion in breast cancer by modulating the miR-1179/TRIM37 axis. Oncol Lett. 2021;22:500.
111. Li J, Kang J, Liu W, et al. Docetaxel-resistant triple-negative breast cancer cell-derived exosomal lncRNA LINC00667 reduces the chemosensitivity of breast cancer cells to docetaxel via targeting miR-200b-3p/Bcl-2 axis. Eur J Histochem. 2022:66.
112. Duan H, Liu Y, Gao Z, Huang W. Recent advances in drug delivery systems for targeting cancer stem cells. Acta Pharm Sin B. 2021;11:55-70.
113. Levine M. Epirubicin in breast cancer: present and future. Clin Breast Cancer. 2000;1 Suppl 1:S62-7.
114. Yao N, Fu Y, Chen L, et al. Long non-coding RNA NONHSAT101069 promotes epirubicin resistance, migration, and invasion of breast cancer cells through NONHSAT101069/miR-129-5p/Twist1 axis. Oncogene. 2019;38:7216-33.
115. Zheng A, Zhang L, Song X, Jin F. Clinical significance of SPRY4-IT1 in efficacy and survival prediction in breast cancer patients undergoing neoadjuvant chemotherapy. Histol Histopathol. 2020;35:361-70.
116. Wang F, Yang S, Lv M, et al. Novel long noncoding RNA 005620 induces epirubicin resistance in triple-negative breast cancer by regulating ITGB1 expression. Front Oncol. 2021;11:592215.
117. Heidelberger C, Chaudhuri NK, Danneberg P, et al. Fluorinated pyrimidines, a new class of tumour-inhibitory compounds. Nature. 1957;179:663-6.
118. Longley DB, Harkin DP, Johnston PG. 5-fluorouracil: mechanisms of action and clinical strategies. Nat Rev Cancer. 2003;3:330-8.
119. Diasio RB, Harris BE. Clinical pharmacology of 5-fluorouracil. Clin Pharmacokinet. 1989;16:215-37.
120. Li X, Wang S, Li Z, et al. The lncRNA NEAT1 facilitates cell growth and invasion via the miR-211/HMGA2 axis in breast cancer. Int J Biol Macromol. 2017;105:346-53.
121. Liang Y, Song X, Li Y, et al. A novel long non-coding RNA-PRLB acts as a tumor promoter through regulating miR-4766-5p/SIRT1 axis in breast cancer. Cell Death Dis. 2018;9:563.
122. Zhang Z, Wang X, Wang Y, et al. Effect of long noncoding RNA CCAT2 on drug sensitivity to 5-fluorouracil of breast cancer cells through microRNA-145 meditated by p53. J Biochem Mol Toxicol. 2022;36:e23176.
123. Luo L, Zhang J, Tang H, et al. LncRNA SNORD3A specifically sensitizes breast cancer cells to 5-FU by sponging miR-185-5p to enhance UMPS expression. Cell Death Dis. 2020;11:329.
124. Yang W, Gu J, Wang X, et al. Inhibition of circular RNA CDR1as increases chemosensitivity of 5-FU-resistant BC cells through up-regulating miR-7. J Cell Mol Med. 2019;23:3166-77.
125. Yao X, Tu Y, Xu Y, Guo Y, Yao F, Zhang X. Endoplasmic reticulum stress confers 5-fluorouracil resistance in breast cancer cell via the GRP78/OCT4/lncRNA MIAT/AKT pathway. Am J Cancer Res. 2020;10:838-55.
126. Villarreal-Garza C, Khalaf D, Bouganim N, et al. Platinum-based chemotherapy in triple-negative advanced breast cancer. Breast Cancer Res Treat. 2014;146:567-72.
127. Galluzzi L, Senovilla L, Vitale I, et al. Molecular mechanisms of cisplatin resistance. Oncogene. 2012;31:1869-83.
128. Kelland L. The resurgence of platinum-based cancer chemotherapy. Nat Rev Cancer. 2007;7:573-84.
129. Rosenberg B, VanCamp L, Trosko JE, Mansour VH. Platinum compounds: a new class of potent antitumour agents. Nature. 1969;222:385-6.
130. Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol. 2014;740:364-78.
131. Siddik ZH. Cisplatin: mode of cytotoxic action and molecular basis of resistance. Oncogene. 2003;22:7265-79.
132. Zhou L, Li H, Sun T, et al. HULC targets the IGF1R-PI3K-AKT axis in trans to promote breast cancer metastasis and cisplatin resistance. Cancer Lett. 2022;548:215861.
133. Su A, Yao K, Zhang H, Wang Y, Zhang H, Tang J. DANCR induces cisplatin resistance of triple-negative breast cancer by KLF5/p27 signaling. Am J Pathol. 2023;193:248-58.
134. García-Venzor A, Mandujano-Tinoco EA, Ruiz-Silvestre A, et al. lncMat2B regulated by severe hypoxia induces cisplatin resistance by increasing DNA damage repair and tumor-initiating population in breast cancer cells. Carcinogenesis. 2020;41:1485-97.
135. Tang T, Cheng Y, She Q, et al. Long non-coding RNA TUG1 sponges miR-197 to enhance cisplatin sensitivity in triple negative breast cancer. Biomed Pharmacother. 2018;107:338-46.
136. Sheng X, Dai H, Du Y, et al. LncRNA CARMN overexpression promotes prognosis and chemosensitivity of triple negative breast cancer via acting as miR143-3p host gene and inhibiting DNA replication. J Exp Clin Cancer Res. 2021;40:205.
137. Wu J, Xu W, Ma L, et al. Formononetin relieves the facilitating effect of lncRNA AFAP1-AS1-miR-195/miR-545 axis on progression and chemo-resistance of triple-negative breast cancer. Aging. 2021;13:18191-222.
138. Wu J, Chen H, Ye M, et al. Long noncoding RNA HCP5 contributes to cisplatin resistance in human triple-negative breast cancer via regulation of PTEN expression. Biomed Pharmacother. 2019;115:108869.
139. Wu B, Yuan Y, Han X, et al. Structure of LINC00511-siRNA-conjugated nanobubbles and improvement of cisplatin sensitivity on triple negative breast cancer. FASEB J. 2020;34:9713-26.
140. Zhou D, Gu J, Wang Y, et al. Long non-coding RNA NEAT1 transported by extracellular vesicles contributes to breast cancer development by sponging microRNA-141-3p and regulating KLF12. Cell Biosci. 2021;11:68.
141. Zhang L, Zhang J, Ni C. Silencing of lncRNA PART1 inhibits proliferation, invasion and migration of breast cancer cells and promotes the efficacy of cisplatin in breast cancer cells. Gen Physiol Biophys. 2020;39:343-54.
142. Zhang M, Yang L, Hou L, Tang X. LncRNA SNHG1 promotes tumor progression and cisplatin resistance through epigenetically silencing miR-381 in breast cancer. Bioengineered. 2021;12:9239-50.
143. Mi H, Wang X, Wang F, et al. SNHG15 contributes To cisplatin resistance in breast cancer through sponging miR-381. Onco Targets Ther. 2020;13:657-66.
144. Zhu M, Xiang H, Peng Z, et al. Silencing the expression of lncRNA SNHG15 may be a novel therapeutic approach in human breast cancer through regulating miR-345-5p. Ann Transl Med. 2022;10:1173.
145. Zhu D, Zhang X, Lin Y, Liang S, Song Z, Dong C. MT1JP inhibits tumorigenesis and enhances cisplatin sensitivity of breast cancer cells through competitively binding to miR-24-3p. Am J Transl Res. 2019;11:245-56.
146. Sella T, Weiss A, Mittendorf EA, et al. Neoadjuvant endocrine therapy in clinical practice: a review. JAMA Oncol. 2021;7:1700-8.
147. Hasson S, Brezis MR, Shachar E, Shachar SS, Wolf I, Sonnenblick A. Adjuvant endocrine therapy in HER2-positive breast cancer patients: systematic review and meta-analysis. ESMO Open. 2021;6:100088.
148. Ballinger TJ, Meier JB, Jansen VM. Current landscape of targeted therapies for hormone-receptor positive, HER2 negative metastatic breast cancer. Front Oncol. 2018;8:308.
149. Hanker AB, Sudhan DR, Arteaga CL. Overcoming endocrine resistance in breast cancer. Cancer Cell. 2020;37:496-513.
150. AlFakeeh A, Brezden-Masley C. Overcoming endocrine resistance in hormone receptor-positive breast cancer. Curr Oncol. 2018;25:S18-27.
152. Barchiesi G, Mazzotta M, Krasniqi E, et al. Neoadjuvant endocrine therapy in breast cancer: current knowledge and future perspectives. Int J Mol Sci. 2020;21:3528.
154. Shi Q, Li Y, Li S, et al. LncRNA DILA1 inhibits Cyclin D1 degradation and contributes to tamoxifen resistance in breast cancer. Nat Commun. 2020;11:5513.
155. Wang J, Xie S, Yang J, et al. The long noncoding RNA H19 promotes tamoxifen resistance in breast cancer via autophagy. J Hematol Oncol. 2019;12:81.
156. Lin X, Dinglin X, Cao S, et al. Enhancer-driven lncRNA BDNF-AS induces endocrine resistance and malignant progression of breast cancer through the RNH1/TRIM21/mTOR cascade. Cell Rep. 2020;31:107753.
157. Xue X, Yang YA, Zhang A, et al. LncRNA HOTAIR enhances ER signaling and confers tamoxifen resistance in breast cancer. Oncogene. 2016;35:2746-55.
158. Li Z, Yu D, Li H, Lv Y, Li S. Long non-coding RNA UCA1 confers tamoxifen resistance in breast cancer endocrinotherapy through regulation of the EZH2/p21 axis and the PI3K/AKT signaling pathway. Int J Oncol. 2019;54:1033-42.
159. Shi YF, Lu H, Wang HB. Downregulated lncRNA ADAMTS9-AS2 in breast cancer enhances tamoxifen resistance by activating microRNA-130a-5p. Eur Rev Med Pharmacol Sci. 2019;23:1563-73.
160. Barazetti JF, Jucoski TS, Carvalho TM, et al. From micro to long: non-coding RNAs in tamoxifen resistance of breast cancer cells. Cancers. 2021;13:3688.
161. Farhan M, Aatif M, Dandawate P, Ahmad A. Non-coding RNAs as mediators of tamoxifen resistance in breast cancers. In: Ahmad A, editor. Breast cancer metastasis and drug resistance. Cham: Springer International Publishing; 2019. pp. 229-41.
162. Yu S, Wang Y, Gong X, et al. LncRNA AGPG confers endocrine resistance in breast cancer by promoting E2F1 activity. Cancer Res. 2023;83:3220-36.
163. Zhang H, Zhang J, Dong L, Ma R. LncRNA ATXN8OS enhances tamoxifen resistance in breast cancer. Open Med. 2021;16:68-80.
164. Horie K, Takagi K, Takeiwa T, et al. Estrogen-inducible LncRNA BNAT1 functions as a modulator for estrogen receptor signaling in endocrine-resistant breast cancer cells. Cells. 2022;11:3610.
165. Liu Y, Li M, Yu H, Piao H. lncRNA CYTOR promotes tamoxifen resistance in breast cancer cells via sponging miR-125a-5p. Int J Mol Med. 2020;45:497-509.
166. Liang X, Zhao Y, Fang Z, et al. DLGAP1-AS2 promotes estrogen receptor signalling and confers tamoxifen resistance in breast cancer. Mol Biol Rep. 2022;49:3939-47.
167. Ma Y, Bu D, Long J, Chai W, Dong J. LncRNA DSCAM-AS1 acts as a sponge of miR-137 to enhance Tamoxifen resistance in breast cancer. J Cell Physiol. 2019;234:2880-94.
168. Ren L, Zhou H, Lei L, Zhang Y, Cai H, Wang X. Long non-coding RNA FOXD3 antisense RNA 1 augments anti-estrogen resistance in breast cancer cells through the microRNA-363/trefoil factor 1/phosphatidylinositol 3-kinase/protein kinase B axis. Bioengineered. 2021;12:5266-78.
169. Li Y, Liu L, Lv Y, et al. Silencing long non-coding RNA HNF1A-AS1 inhibits growth and resistance to TAM of breast cancer cells via the microRNA-363/SERTAD3 axis. J Drug Target. 2021;29:742-53.
170. Kim CY, Oh JH, Lee JY, Kim MH. The LncRNA HOTAIRM1 promotes tamoxifen resistance by mediating HOXA1 expression in ER+ breast cancer cells. J Cancer. 2020;11:3416-23.
171. Lee J, Kang B, Kim EA, et al. Differential expression of long non-coding RNA IGF2-as in tamoxifen-resistant breast cancer cells. Biomedicines. 2025;13:2087.
172. Peng WX, Koirala P, Zhou H, et al. Lnc-DC promotes estrogen independent growth and tamoxifen resistance in breast cancer. Cell Death Dis. 2021;12:1000.
173. Liao M, Webster J, Coonrod EM, Weilbaecher KN, Maher CA, White NM. BCAR4 expression as a predictive biomarker for endocrine therapy resistance in breast cancer. Clin Breast Cancer. 2024;24:368-75.e2.
174. Chen X, Ding JC, Hu GS, et al. Estrogen-induced LncRNA, LINC02568, promotes estrogen receptor-positive breast cancer development and drug resistance through both in trans and in cis mechanisms. Adv Sci. 2023;10:e2206663.
175. Ma T, Liang Y, Li Y, et al. LncRNA LINP1 confers tamoxifen resistance and negatively regulated by ER signaling in breast cancer. Cell Signal. 2020;68:109536.
176. Sun W, Xu X, Jiang Y, et al. Transcriptome analysis of luminal breast cancer reveals a role for LOL in tumor progression and tamoxifen resistance. Int J Cancer. 2019;145:842-56.
177. Feng J, Wen T, Li Z, et al. Cross-talk between the ER pathway and the lncRNA MAFG-AS1/miR-339-5p/ CDK2 axis promotes progression of ER+ breast cancer and confers tamoxifen resistance. Aging. 2020;12:20658-83.
178. Wu C, Sun X, Lu Y, et al. Cancer-associated fibroblast promotes tamoxifen resistance in estrogen receptor positive breast cancer via exosomal LncRNA PRKCQ-AS1/miR-200a-3p/MKP1 axis-mediated apoptosis suppression. J Exp Clin Cancer Res. 2025;44:274.
179. Peng WX, Huang JG, Yang L, Gong AH, Mo YY. Linc-RoR promotes MAPK/ERK signaling and confers estrogen-independent growth of breast cancer. Mol Cancer. 2017;16:161.
180. Khan MI, Ahmad A. LncRNA SNHG6 sponges miR-101 and induces tamoxifen resistance in breast cancer cells through induction of EMT. Front Oncol. 2022;12:1015428.
181. Fang J, Li K, Huang C, Xue H, Ni Q. LncRNA TTN-AS1 confers tamoxifen resistance in breast cancer via sponging miR-107 to modulate PI3K/AKT signaling pathway. Am J Transl Res. 2022;14:2267-79.
182. Ratre P, Mishra K, Dubey A, Vyas A, Jain A, Thareja S. Aromatase inhibitors for the treatment of breast cancer: a journey from the scratch. Anticancer Agents Med Chem. 2020;20:1994-2004.
183. Sood A, Lang DK, Kaur R, Saini B, Arora S. Relevance of aromatase inhibitors in breast cancer treatment. Curr Top Med Chem. 2021;21:1319-36.
184. Nagini S. Breast cancer: current molecular therapeutic targets and new players. Anticancer Agents Med Chem. 2017;17:152-63.
185. Chen X, Luo R, Zhang Y, et al. Long noncoding RNA DIO3OS induces glycolytic-dominant metabolic reprogramming to promote aromatase inhibitor resistance in breast cancer. Nat Commun. 2022;13:7160.
186. Huang S, Chi Y, Chi W, et al. LINC00309 is associated with short disease-free survival in breast cancer. Cancer Cell Int. 2019;19:210.
187. Ingle JN, Xie F, Ellis MJ, et al. Genetic polymorphisms in the long noncoding RNA MIR2052HG offer a pharmacogenomic basis for the response of breast cancer patients to aromatase inhibitor therapy. Cancer Res. 2016;76:7012-23.
188. Gu G, Dustin D, Fuqua SA. Targeted therapy for breast cancer and molecular mechanisms of resistance to treatment. Curr Opin Pharmacol. 2016;31:97-103.
189. Schlam I, Tarantino P, Morganti S, et al. Emerging targeted therapies for early breast cancer. Drugs. 2022;82:1437-51.
190. Lau KH, Tan AM, Shi Y. New and emerging targeted therapies for advanced breast cancer. Int J Mol Sci. 2022;23:2288.
191. Greenwalt I, Zaza N, Das S, Li BD. Precision medicine and targeted therapies in breast cancer. Surg Oncol Clin N Am. 2020;29:51-62.
192. Jacobs AT, Martinez Castaneda-Cruz D, Rose MM, Connelly L. Targeted therapy for breast cancer: an overview of drug classes and outcomes. Biochem Pharmacol. 2022;204:115209.
193. Clarke R, Tyson JJ, Dixon JM. Endocrine resistance in breast cancer-an overview and update. Mol Cell Endocrinol. 2015;418 Pt 3:220-34.
195. Curigliano G, Criscitiello C. Successes and limitations of targeted cancer therapy in breast cancer. In: Peters S, Stahel R, editors. Successes and limitations of targeted cancer therapy; 2014. pp. 15-35.
196. Henriques B, Mendes F, Martins D. Immunotherapy in breast cancer: when, how, and what challenges? Biomedicines. 2021;9:1687.
197. Wang ZH, Zheng ZQ, Jia SC, et al. Trastuzumab resistance in HER2-positive breast cancer: mechanisms, emerging biomarkers and targeting agents. Front Oncol. 2022;12:1006429.
198. Ghafouri-Fard S, Tamizkar KH, Hussen BM, Taheri M. An update on the role of long non-coding RNAs in the pathogenesis of breast cancer. Pathol Res Pract. 2021;219:153373.
199. Dong H, Wang W, Mo S, et al. Long non-coding RNA SNHG14 induces trastuzumab resistance of breast cancer via regulating PABPC1 expression through H3K27 acetylation. J Cell Mol Med. 2018;22:4935-47.
200. Zhu HY, Bai WD, Ye XM, Yang AG, Jia LT. Long non-coding RNA UCA1 desensitizes breast cancer cells to trastuzumab by impeding miR-18a repression of Yes-associated protein 1. Biochem Biophys Res Commun. 2018;496:1308-13.
201. Ghazimoradi MH, Babashah S. The role of CircRNA/miRNA/mRNA axis in breast cancer drug resistance. Front Oncol. 2022;12:966083.
202. Zou Y, Yang A, Chen B, et al. crVDAC3 alleviates ferroptosis by impeding HSPB1 ubiquitination and confers trastuzumab deruxtecan resistance in HER2-low breast cancer. Drug Resist Updat. 2024;77:101126.
203. Misir S, Yaman SO, Petrović N, Sumer C, Hepokur C, Aliyazicioglu Y. circRNAs in drug resistance of breast cancer. Oncol Res. 2022;30:157-72.
204. Guo J, Li K, Ming Y, et al. A circular RNA overcomes acquired resistance to BET inhibitors by antagonizing IGF2BP2-mediated c-MYC translation in TNBC. Proc Natl Acad Sci USA. 2025;122:e2504320122.
205. Shi SJ, Wang LJ, Yu B, Li YH, Jin Y, Bai XZ. LncRNA-ATB promotes trastuzumab resistance and invasion-metastasis cascade in breast cancer. Oncotarget. 2015;6:11652-63.
206. Song X, Liu S, Zeng Y, Cai Y, Luo H. BANCR-containing extracellular vesicles enhance breast cancer resistance. J Biol Chem. 2025;301:108304.
207. Ye X, Liu Q, Qin X, et al. BCAR4 facilitates trastuzumab resistance and EMT in breast cancer via sponging miR-665 and interacting with YAP1. FASEB J. 2024;38:e23589.
208. Sun Z, Zhang C, Wang T, Shi P, Tian X, Guo Y. Correlation between long non-coding RNAs (lncRNAs) H19 expression and trastuzumab resistance in breast cancer. J Cancer Res Ther. 2019;15:933-40.
209. Chen T, Liu Z, Zeng W, Huang T. Down-regulation of long non-coding RNA HOTAIR sensitizes breast cancer to trastuzumab. Sci Rep. 2019;9:19881.
210. Bai W, Peng H, Zhang J, et al. LINC00589-dominated ceRNA networks regulate multiple chemoresistance and cancer stem cell-like properties in HER2+ breast cancer. NPJ Breast Cancer. 2022;8:115.
211. Liu C, Lu C, Yixi L, et al. Exosomal Linc00969 induces trastuzumab resistance in breast cancer by increasing HER-2 protein expression and mRNA stability by binding to HUR. Breast Cancer Res. 2023;25:124.
212. Yu Q, Li Y, Peng S, Li J, Qin X. Exosomal-mediated transfer of OIP5-AS1 enhanced cell chemoresistance to trastuzumab in breast cancer via up-regulating HMGB3 by sponging miR-381-3p. Open Med. 2021;16:512-25.
213. Han M, Qian X, Cao H, et al. lncRNA ZNF649-AS1 induces trastuzumab resistance by promoting ATG5 expression and autophagy. Mol Ther. 2020;28:2488-502.
214. Dong H, Han J, Chen X, Sun H, Han M, Wang W. LncRNA ZNF649-AS1 promotes trastuzumab resistance and TAM-dependent PD-L1 expression in breast cancer by regulating EXOC7 alternative splicing. Arch Biochem Biophys. 2024;761:110128.
215. Li W, Zhai L, Wang H, et al. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget. 2016;7:27778-86.
216. Convit J, Montesinos H, Oviedo H, et al. Autologous tumor lysate/Bacillus Calmette-Guérin immunotherapy as an adjuvant to conventional breast cancer therapy. Clin Transl Oncol. 2015;17:884-7.
217. Morales MA, Barrera Rodríguez R, Santiago Cruz JR, Teran LM. Overview of new treatments with immunotherapy for breast cancer and a proposal of a combination therapy. Molecules. 2020;25:5686.
218. Pilipow K, Darwich A, Losurdo A. T-cell-based breast cancer immunotherapy. Semin Cancer Biol. 2021;72:90-101.
219. Schmid P, Adams S, Rugo HS, et al. Atezolizumab and Nab-paclitaxel in advanced triple-negative breast cancer. N Engl J Med. 2018;379:2108-21.
220. Jia H, Truica CI, Wang B, et al. Immunotherapy for triple-negative breast cancer: existing challenges and exciting prospects. Drug Resist Updat. 2017;32:1-15.
221. Li S, Sun X, Li J, et al. A novel prognostic signature of immune-related long noncoding RNA pairs for tumor-infiltrating immune cells and drug susceptibility in breast cancer. DNA Cell Biol. 2022;41:103-15.
222. Shen S, Chen X, Hu X, Huo J, Luo L, Zhou X. Predicting the immune landscape of invasive breast carcinoma based on the novel signature of immune-related lncRNA. Cancer Med. 2021;10:6561-75.
223. Xu Y, Zheng Q, Zhou T, Ye B, Xu Q, Meng X. Necroptosis-related LncRNAs signature and subtypes for predicting prognosis and revealing the immune microenvironment in breast cancer. Front Oncol. 2022;12:887318.
224. Zhang X, Zhang X, Li G, et al. A novel necroptosis-associated lncRNA signature can impact the immune status and predict the outcome of breast cancer. J Immunol Res. 2022;2022:3143511.
225. Zhang D, Lu W, Zhuo Z, Wang Y, Zhang W, Zhang M. Comprehensive analysis of a cuproptosis-related ceRNA network implicates a potential endocrine therapy resistance mechanism in ER-positive breast cancer. BMC Med Genomics. 2023;16:96.
226. Liao B, Wang J, Xie Y, Luo H, Min J. LINK-A: unveiling its functional role and clinical significance in human tumors. Front Cell Dev Biol. 2024;12:1354726.
227. Hu Q, Ye Y, Chan LC, et al. Oncogenic lncRNA downregulates cancer cell antigen presentation and intrinsic tumor suppression. Nat Immunol. 2019;20:835-51.
228. Lian B, Li J, Tang S, Li T, Li J. Targeting LINC02544/miR-497-5p/CAPRIN1 axis via exosome-based siRNA to overcome immunotherapy resistance in triple-negative breast cancer. Mol Med. 2025;31:278.
229. Sun C, Ye Y, Tan Z, et al. Tumor-associated nonmyelinating Schwann cell-expressed PVT1 promotes pancreatic cancer kynurenine pathway and tumor immune exclusion. Sci Adv. 2023;9:eadd6995.
230. Famta P, Shah S, Khatri DK, Guru SK, Singh SB, Srivastava S. Enigmatic role of exosomes in breast cancer progression and therapy. Life Sci. 2022;289:120210.
231. Rezaee M, Mohammadi F, Keshavarzmotamed A, et al. The landscape of exosomal non-coding RNAs in breast cancer drug resistance, focusing on underlying molecular mechanisms. Front Pharmacol. 2023;14:1152672.
232. Huang D, Chen J, Yang L, et al. NKILA lncRNA promotes tumor immune evasion by sensitizing T cells to activation-induced cell death. Nat Immunol. 2018;19:1112-25.
233. Pei X, Wang X, Li H. LncRNA SNHG1 regulates the differentiation of Treg cells and affects the immune escape of breast cancer via regulating miR-448/IDO. Int J Biol Macromol. 2018;118:24-30.
234. Yang H, Qi C, Li B, Cheng L. Non-coding RNAs as novel biomarkers in cancer drug resistance. Curr Med Chem. 2022;29:837-48.
235. Wu J, Zhang C, Li H, Zhang S, Chen J, Qin L. Competing endogenous RNAs network dysregulation in oral cancer: a multifaceted perspective on crosstalk and competition. Cancer Cell Int. 2024;24:431.
236. To KKW, Zhang H, Cho WC. Competing endogenous RNAs (ceRNAs) and drug resistance to cancer therapy. Cancer Drug Resist. 2024;7:37.
237. Wu B, Zhang Y, Yu Y, et al. Long noncoding RNA H19: a novel therapeutic target emerging in oncology via regulating oncogenic signaling pathways. Front Cell Dev Biol. 2021;9:796740.
238. Su WY, Tian LY, Guo LP, Huang LQ, Gao WY. PI3K signaling-regulated metabolic reprogramming: from mechanism to application. Biochim Biophys Acta Rev Cancer. 2023;1878:188952.
239. Cirone M. Cancer cells dysregulate PI3K/AKT/mTOR pathway activation to ensure their survival and proliferation: mimicking them is a smart strategy of gammaherpesviruses. Crit Rev Biochem Mol Biol. 2021;56:500-9.
240. Raju GSR, Pavitra E, Bandaru SS, et al. HOTAIR: a potential metastatic, drug-resistant and prognostic regulator of breast cancer. Mol Cancer. 2023;22:65.
241. Barwal TS, Sharma U, Bazala S, et al. MicroRNAs and long noncoding RNAs as novel therapeutic targets in estrogen receptor-positive breast and ovarian cancers. Int J Mol Sci. 2021;22:4072.
242. Hu X, Zhang Q, Xing W, Wang W. Role of microRNA/lncRNA intertwined with the Wnt/β-Catenin axis in regulating the pathogenesis of triple-negative breast cancer. Front Pharmacol. 2022;13:814971.
243. Ebrahimnezhad M, Asl SH, Rezaie M, Molavand M, Yousefi B, Majidinia M. lncRNAs: new players of cancer drug resistance via targeting ABC transporters. IUBMB Life. 2024;76:883-921.
244. Rembiałkowska N, Kocik Z, Kłosińska A, et al. Inflammation-driven genomic instability: a pathway to cancer development and therapy resistance. Pharmaceuticals. 2025;18:1406.
245. Han C, Zhang C, Wang H, Zhao L. Exosome-mediated communication between tumor cells and tumor-associated macrophages: implications for tumor microenvironment. Oncoimmunology. 2021;10:1887552.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.